La زانو یکی از پیچیده ترین مفصل های بدن انسان است که شامل استخوان ران یا استخوان ران، استخوان ران یا استخوان ران و استخوان ران و زانو، یا پاتلا، در میان دیگر بافت های نرم است. سینه ها استخوان ها را به عضلات متصل می کنند در حالی که رباط ها استخوان های مفصل زانو را متصل می کنند. دو قطعه گوه شکل غضروف، شناخته شده به عنوان منیسک، پایداری را به مفصل زانو ارائه می کنند. هدف مقاله زیر نشان دادن و همچنین بحث در مورد آناتومی مفصل زانو و بافت نرم اطراف آن است.
فهرست
چکیده
- زمینه: اطلاعات مربوط به ساختار ، ترکیب و عملکرد منیسک زانو در چندین منبع و زمینه پراکنده شده است. این بررسی شامل شرح مختصر و مفصلی از منیسک زانو شامل آناتومی ، ریشه شناسی ، فیلوژنی ، فراساختار و بیوشیمی ، آناتومی عروقی و نورواناتومی ، عملکرد بیومکانیکی ، بلوغ و پیری و روش های تصویربرداری است.
- اکتساب شواهد: جستجوی ادبیات توسط بررسی مقالات PubMed و OVID منتشر شده از 1858 به 2011 انجام شد.
- نتایج: این مطالعه ویژگی های ساختاری، ترکیبات و عملکردی منیسکی را که ممکن است مربوط به ارائه بالینی، تشخیص و تعمیرات جراحی باشد، مشخص می کند.
- نتیجه گیری: درک آناتومی عادی و بیومکانیک منیسیک پیش نیاز لازم برای درک پاتوژنز اختلالات مربوط به زانو است.
- کلید واژه ها: زانو، منیسک، آناتومی، عملکرد
معرفی
هنگامی که به عنوان یک بقایای جنینی بدون عملکرد توصیف می شود ، اکنون 162 مینیسک برای عملکرد طبیعی و سلامت طولانی مدت مفصل زانو حیاتی شناخته شده است. men مینیسک ها باعث افزایش ثبات برای بیان مفصل ران ، توزیع بار محوری ، جذب شوک و ایجاد روغنکاری می شوند. و تغذیه مفصل زانو .4,91,152,153،XNUMX،XNUMX،XNUMX
آسیب های منیسکی به عنوان یک علت عوارض اسکلتی عضلانی شناخته شده است. ساختار منحصر به فرد و پیچیده Menisci درمان و تعمیر برای بیمار، جراح و فیزیوتراپیست را به چالش کشیدن. علاوه بر این آسیب درازمدت ممکن است منجر به تغییرات مفصلی دژنراتیو مانند ایجاد osteophyte، دژنراسیون مفصلی غضروف، تنگ شدن فضای مفصلی و استئوآرتریت علامتدار شود. 36,45,92 حفظ منیسکی بستگی به حفظ ترکیب و سازماندهی متمایز آن دارد.
آناتومی Menisci
ریشهشناسی منسکی
کلمه meniscus از کلمه یونانی m؟ niskos ، به معنای هلال ، کوچک کننده m؟ n ؟، به معنای ماه است. oon
فیزیولوژی انسانی و آناتومی تطبیقی
هومیندها ویژگی های آناتومیکی و عملکردی مشابهی دارند، از جمله یک femur ديستال bicondylar، رباط صلیبی داخل مفصلی، منیسکی و غیرهثباتی. xNUMX این ویژگی های مورفولوژیکی مشابه، منعکس کننده یک ژنتیک مشترک ژنتیکی است که می تواند بیش از 40,66 میلیون سال ردیابی شود. 300
در اصل و نسب پستانداران که منجر به انسان می شود ، انسان کشی ها تقریباً 3 تا 4 میلیون سال پیش به حالت دو پا تبدیل می شوند و 1.3 میلیون سال پیش ، مفصل مدرن کشکککویی (با وجه کشکک جانبی طولانی تر و مطابق با استخوان ران استخوان ران جانبی) ایجاد شد. 164 Tardieu انتقال از دو پله پالیزم گاه به گاه به دو پدالیسم دائمی را مورد بررسی قرار داد و مشاهده کرد که پستانداران شامل یک مینیسک فیبروکارتیلاژینوس داخلی و جانبی هستند ، با اینکه مینیسک داخلی از نظر ریخت شناسی در همه نخستی های اولیه (هلال با 2 درج استخوان درشت نی شکل) مشابه است. 163 در مقابل ، مینیسک جانبی مشاهده شد شکل متغیرتر باشد در Homo sapiens وجود 2 محل وارد آمدن استخوان درشت نی 1 قدامی و 1 خلفی نشانگر یک عمل عادی از حرکات کششی کامل مفصل زانو در طی مراحل ایستادن و چرخش راه رفتن دو پا است. 20,134,142,163,168،XNUMX،XNUMX،XNUMX،XNUMX
جنین شناسی و توسعه
شکل مشخصی از منیسک جانبی و مزایای بین هفته های 8 و 10th از زمان بارداری به دست می آید. 53,60 آنها از یک تراکم لایه میان بافت مزانشیمال به منظور تشکیل پیوستگی به کپسول متصل به اطراف آن بوجود می آیند. 31,87,110 Menisci در حال رشد بسیار سلولی است و عروق، با خونریزی وارد شده از حاشیه و گسترش از طریق تمام عرض menisci.31 همانطور که جنین همچنان به توسعه، کاهش تدریجی سلولیت منیسیک با افزایش همراه با محتوای کلاژن در یک محدوده تنظیم حرکت. 30,31 حرکت مشترک و تنش پس از زایمان وزن تنشی عوامل مهمی در تعیین جهت گیری فیبر کلاژن هستند. با بزرگسالی، تنها 10٪ محیطی به 30٪ دارای خون است. 12,31
علیرغم این تغییرات بافت شناسی، نسبت فلج تیبالی پوشیده شده توسط منیسک های مربوطه نسبتا ثابت است در طول توسعه جنین، با منیسکی میانی و جانبی حدود 60٪ و 80٪ از مناطق سطح به ترتیب. 31
آناتومی ناخالص
بررسی کامل منیسک زانو یک بافت صاف و روان کننده را نشان می دهد (شکل 1). آنها قواي فيبروتيكه اي شكافي هستند كه در جنبه هاي مياني و جانبي مفصل زانو قرار دارند (شکل 2A). مرز محیطی، عروقی (همچنین به عنوان منطقه قرمز) هر منیسک ضخیم، محدب و متصل به کپسول مفصلی است. مرز درونی (همچنین به عنوان منطقه سفید شناخته می شود) به یک لبه آزاد نازک متصل می شود. سطوح برتر منیسک ها دارای مقعر هستند، که می تواند مفصل موثر با فتق های فمورال محدب مربوطه را ایجاد کند. سطوح پایین تر برای تطابق با فلات تیبیس (شکل 1) مسطح است. 28,175
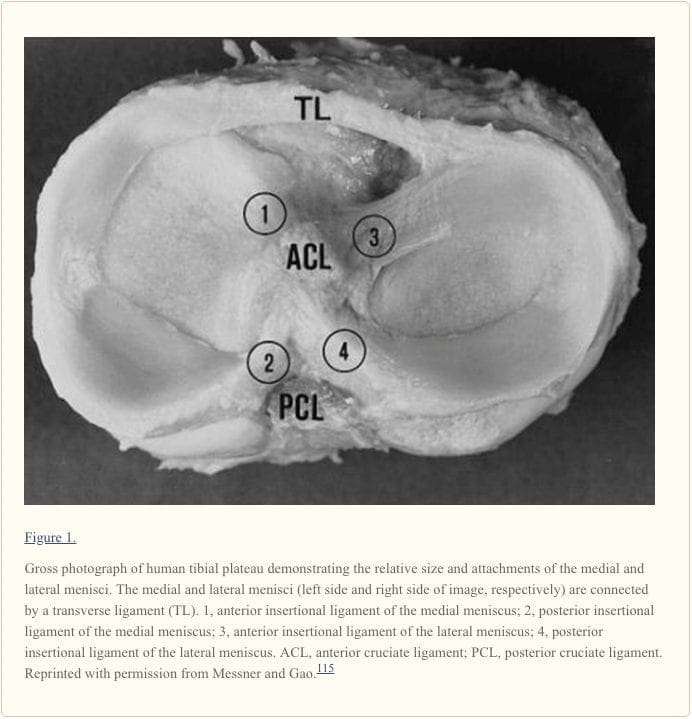
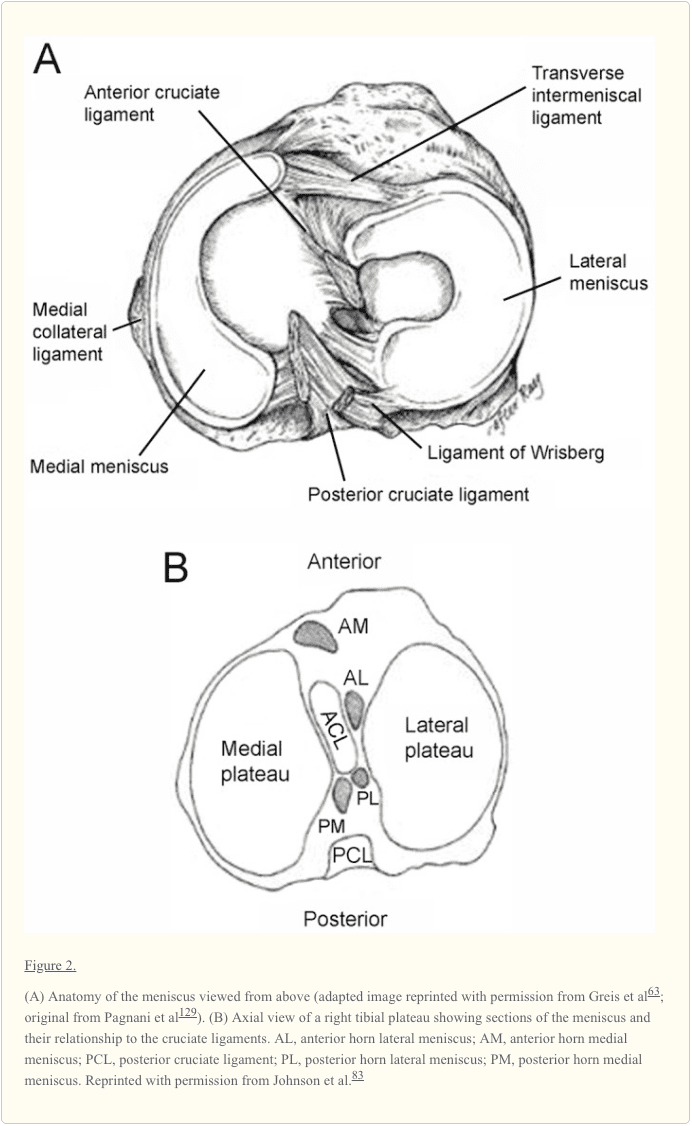
منیسک مدیونی منیسک محیطی نیمه مدور تقریبا 35 میلی متر در قطر (قدام به عقب) اندازه گیری می شود و به طور قابل توجهی گسترده تر از عقب است. 175 شاخ قدامی به فلات تیبیا در نزدیکی فسکت اینترچندیلار قدام به رباط متقاطع قدامی (ACL) متصل است. تغییر مکان قابل توجهی در محل دلبستگی شاخ قدامی منیسک مدیا وجود دارد. شاخ خلفی به مجرای خلفی اینترکندیلار تیبیا بین منیسک جانبی و رباط صلیبی خلفی (PCL؛ شکل 1 و 2B) 2B متصل است. جانسون و همکاران، سایت های مری و مینیسک و روابط توپوگرافی خود را با اشیای اطراف آناتومیک زانو بررسی کردند. 82 آنها دریافتند که سایت های ورودی قدامی و خلفی منیسک متوسط بیشتر از منیسک جانبی هستند. محدوده محل قرارگیری شاخ مفاصل منیسک مزوتراپی بزرگتر بود، اندازه گیری 61.4 mm2، در حالی که شاخ خلفی منیسک جانبی کمترین بود، در 28.5 mm2.82
قسمت تیبیال اتصال کپسولی رباط کرونری است. در نقطه میانی، منیسک داخلی از طریق تراکم در کپسول مفصلی به نام رباط جانبی داخلی عمیق، محکمتر به استخوان ران میچسبد. 175 رباط عرضی یا «بینمنیسکال» یک نوار فیبری از بافت است که شاخ قدامی را به هم متصل میکند. از منیسک داخلی به شاخ قدامی منیسک جانبی (شکل 1 و and2A2A).
منیسک جانبی منیسک جانبی تقریبا دایره ای است با عرض تقریبا یکنواخت از قدامی تا خلفی (شکل 1 و and2A) .2A). آن قسمت بزرگتر (~ 80٪) از سطح مفصلی را نسبت به منیسک مدیایی (~ 60٪) اشغال می کند و بیشتر همراه است. 10,31,165 هر دو شاخۀ منیسک جانبی به تیبیا متصل می شوند. جایگزینی شاخ قدامی منیسک جانبی در قسمت قدامی بین سرخجه بین کاندیدال و در مجاورت محل اتصال گسترده ACL (شکل 2B) قرار دارد. 9,83 شاخ خلفی منیسک جانبی بعد از عقب به ستون فقرات تیبيال و فقط قدامي قرار دادن شاخ خلفی منیسک مدفوع (شکل 2B) .83 منیسک جانبی به آرامی به رباط کپسول متصل می شود؛ با این حال، این الیاف به رباط جانبی جانبی اضافه نمی کنند. شاخ خلفی منیسک جانبی به جنبه داخلی کاندیل فمورال میدلی از طریق رباط های منسوفومورال قدامی و خلفی هامفری و وریسبرگ متصل می شود که در نزدیکی منبع PCL قرار دارند (شکل 1 و and22) .75
پیوند منیسوفومورال. ادبیات ناسازگاری قابل توجهی را در حضور و اندازه رباط های منیسکوفمورال منیسک جانبی گزارش می کند. ممکن است هیچ کدام وجود نداشته باشد، 1، 2، یا 4.؟ در صورت وجود، این رباط های جانبی از شاخ خلفی منیسک جانبی به سمت جانبی کندیل داخلی فمورال عرضی می کنند. آنها بلافاصله در مجاورت ضمیمه فمورال PCL قرار می گیرند (شکل 1 و و22).
در یک سری مطالعات Harner و همکاران، سطح مقطع لیگامان را اندازه گیری کردند و دریافتند که لنز منیسوفومورال به طور متوسط 20٪ از اندازه PCL (محدوده 7٪ -35٪) است. 69,70 با این حال، اندازه منطقه مستقل تنها بدون دانستن زاویه قرارگیری یا تراکم کلاژن، قدرت نسبی آنها را نشان نمی دهد. 115 عملکرد این رباط ها هنوز ناشناخته مانده است. آنها ممکن است شاخ خلفی منیسک جانبی را در جهت قدامی افزایش دهند تا منافذ مناسکوتیوبال و لگن جانبی فمورال را افزایش دهند. 75
فراصوت و بیوشیمی
ماتریکس خارج سلولی
منیسک یک ماتریس خارج سلولی متراکم (ECM) است که در درجه اول از آب (72٪) و کلاژن (22٪) تشکیل شده و با سلولها در میان است. 9,55,56,77،XNUMX،XNUMX،XNUMX پروتئوگلیکان ها ، پروتئین های غیر کلاژن و گلیکوپروتئین ها وزن خشک باقی مانده را تشکیل می دهند. cells سلولهای مینیسک ECM را که خصوصیات مواد بافت را تعیین می کند ، سنتز و حفظ می کنند.
سلولهای منیسکی به عنوان فیبروچاندروسیت ها نامیده می شوند، زیرا آنها به نظر می رسد ترکیبی از فیبروبلاست ها و کندروسیت ها هستند. 111,177 سلول های لایه سطحی سطحی منیسکی فوزیک شکل یا اسپیندل (بیشتر فیبروبلاستی) هستند، در حالی که سلول ها عمیق تر در منیسک ها تخم مرغی یا چند ضلعی هستند (بیشتر از نوع کاندروسیتیک). مورفولوژی سلول 55,56,178 بین مکان های محیطی و مرکزی در menisci متفاوت نیست. 56
هر دو نوع سلول شامل سلول های اندوپلاسمی فراوان و پیچیدگی گلژی هستند. میتوکندریا تنها گاهی تجسم می شود، که نشان می دهد راه اصلی برای تولید انرژی فیبروکندروسیت ها در محیط اتواسکلریت احتمالا گلیکولیز بی هوازی است. 112
آب
در منیسک طبیعی سالم، مایع بافت 65٪ تا 70٪ از وزن کل را نشان می دهد. اکثر آب درون بافت در حوزه های پروتئول کلین حلال نگهداری می شود. محتوای آب بافت منیسک در مناطق خلفی بیشتر از مناطق مرکزی یا قدامی است. نمونه های بافت از سطوح سطحی و عمیق محتوای مشابهی داشتند. 135
فشارهای هیدرولیک بزرگ برای غلبه بر کشیدن مقاومت اصطکاکی در جریان جریان مایع از طریق بافت منیسک مورد نیاز است. بنابراین، تعاملات بین آب و چارچوب ماکرومولکولی ماتریکس به طور قابل توجهی بر خواص ویسکولا الاستیک از بافت تاثیر می گذارد.
کلاژن
Collagens در درجه اول مسئول قدرت کشش منیسک هستند؛ آنها به 75٪ از وزن خشک ECM.77 کمک می کنند ECM به طور عمده از کلاژن نوع I (90٪ خشک وزن) با مقادیر متغیر از انواع II، III، V، و VI تشکیل شده است. 43,44,80,112,181 غلظت نوع I کلاژن فیبروکواسترویید منیسک را از مفصلی (هیالین) غضروف متمایز می کند. کولاژن ها به شدت توسط هیدروکسی پالیدین آلومینیوم متصل شده اند. 44
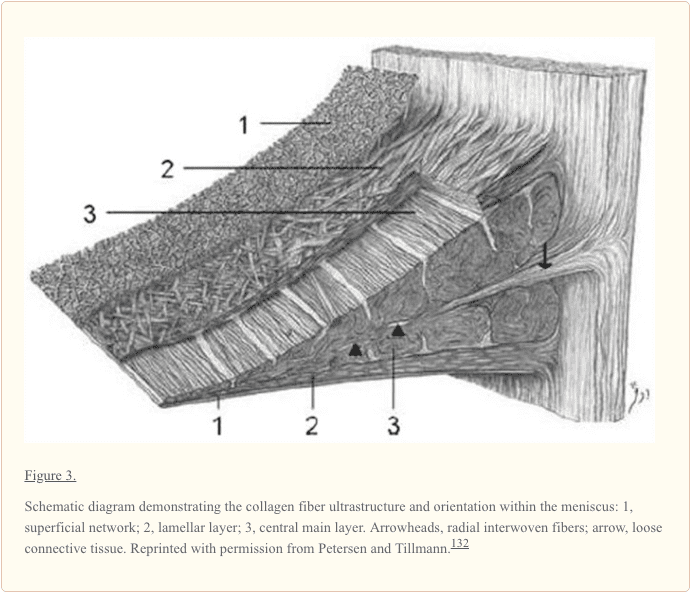
آرایش فیبر کلاژن برای انتقال یک بار فشاری عمودی به تنشهای حلقهای محیطی ایدهآل است (شکل 3). الیاف کلاژن نوع I به صورت محیطی در لایههای عمیقتر منیسک، موازی با مرز محیطی جهتگیری میکنند. این فیبرها اتصالات لیگامانی شاخ های منیسک را با سطح مفصلی تیبیا مخلوط می کنند (شکل 57). الیاف "پیوند" جهت شعاعی نیز در ناحیه عمیق وجود دارند و برای ایجاد یکپارچگی ساختاری بین الیاف محیطی پراکنده یا بافته می شوند (شکل 3). حاوی بلورهای بلند و باریک از فسفر، کلسیم و منیزیم در تجزیه و تحلیل رونتژنوگرافی با پروب الکترونی است.
پروتئین های ماتریکس غیر کلاژانیک، مانند فیبرنرکتین، از 8٪ تا 13 درصد وزن خشک ارگانیک استفاده می کنند. فیبرنکتین در بسیاری از فرآیندهای سلولی، از جمله تعمیر بافت، جنین زایی، لخته شدن خون و مهاجرت / چسبندگی سلولی نقش دارد. الاستین کمتر از 0.6 درصد وزن خشک منیسک را تشکیل می دهد. محلی سازی اشعه ماوراء بنفش روشن نیست. این احتمالا به طور مستقیم با کلاژن تعامل دارد تا سبب انعطاف پذیری در بافت شود. **
پروتئوگلیکان
پروژوسیت ها در داخل یک شبکه خوب از فیبرهای کلاژن قرار دارند و مولکول های هیدروفیلی بزرگ و مولد هیدروفیلی هستند که منعکس می شوند و 1٪ به 2٪ از وزن خشک می پردازند. 58 آنها توسط یک پروتئین هسته با 1 یا بیشتر زنجیره ای از گلیکوزامینوگلیکان متصل به کوالانسان (شکل 4) تشکیل می شوند. 122 اندازه این مولکول ها بوسیله تعامل خاص با اسید هیالورونیک افزایش می یابد. 67,72 مقدار پروتئگلیکان ها در منیسک یک هشتم از غضروف مفصلی 2,3 است و ممکن است بسته به محل نمونه و سن بیمار. 49
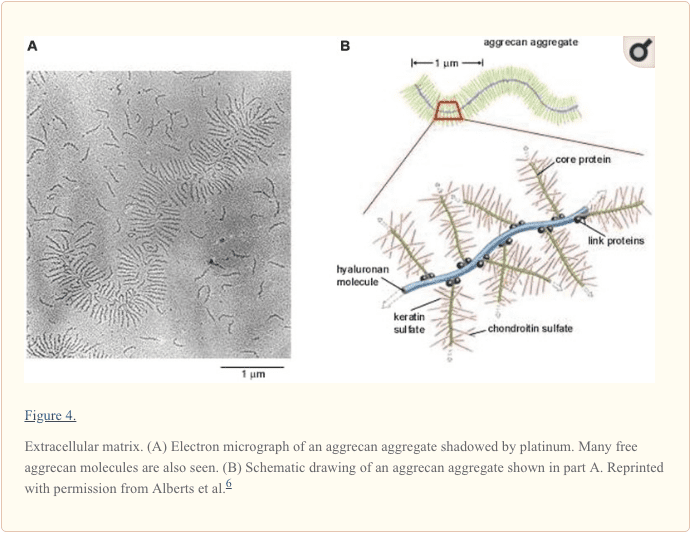
پروتئوگلیکان ها در ECM به دلیل ساختار تخصصی ، تراکم بار ثابت بالا و نیروهای دافعه بار شارژ ، مسئول هیدراتاسیون هستند و ظرفیت بالایی را در بافت برای مقاومت در برابر فشارهای فشاری فراهم می کنند. profile مشخصات گلیکوزآمینوگلیکان انسان بالغ طبیعی مینیسک شامل کندرویتین-6-سولفات (40٪) ، کندرویتین-4-سولفات (10٪ تا 20٪) ، درماتان سولفات (20٪ تا 30٪) و کراتین سولفات (15٪ ؛ شکل 4) است .65,77,99,159،58,77 ، XNUMX،XNUMX بالاترین غلظت گلیکوزآمینوگلیکان در شاخهای مینیسک و نیمه داخلی مینیسک ها در مناطق اصلی تحمل وزن وجود دارد. XNUMX،XNUMX
Aggrecan پروتئگلیکان اصلی در منیسک های انسانی است و عمدتا مسئولیت خواص فشرده سازی ویسکوزالاستیک خود را دارد (شکل 5). پروتئولگلانهای کوچکتر مانند دکورین، biglycan و فیبرومدولین در مقادیر کمتری یافت میشوند. 124,151 هگزازامین به 1٪ به وزن خشک ECM.57,74 عمل می کند. عملکرد دقیق هر یک از این پروتئول کلین ها در منیسک هنوز کاملا مشخص نشده است.
ماتریکس گلیکوپروتئین ها
غضروف منیسک حاوی طیف وسیعی از گلیکوپروتئین های ماتریکس است که هویت و عملکرد آنها هنوز مشخص نشده است. الکتروفورز و رنگآمیزی بعدی ژلهای پلی آکریل آمید نوارهایی با وزن مولکولی متفاوت از چند کیلو دالتون تا بیش از 200 کیلو دالتون را نشان میدهد. این پروتئین در ماتریکس به شکل کمپلکس متصل به دی سولفید با وزن مولکولی بالا قرار دارد.
گلیکوپروتئین های چسبنده یک زیرگروه از گلیکوپروتئین های ماتریکس هستند. این ماکرومولکولها تا حدی مسئول پیوند با مولکولهای ماتریکس و / یا سلولها هستند. بنابراین مولکول های چسبندگی بین مولکولی به عنوان یکی از اجزای مهم در ساختار supramolecular از مولکول های خارج سلولی منیسک هستند. 150 سه مولکول درون منیسک شناسایی شده اند: نوع کلاژن VI، فیبرونکتین و ترومبوسپوندین. 112,118,181
آناتومی عروقی
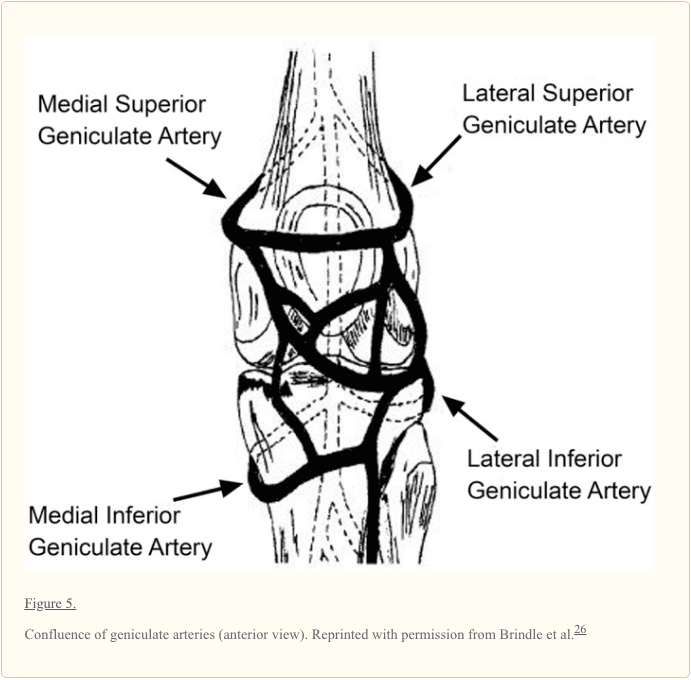
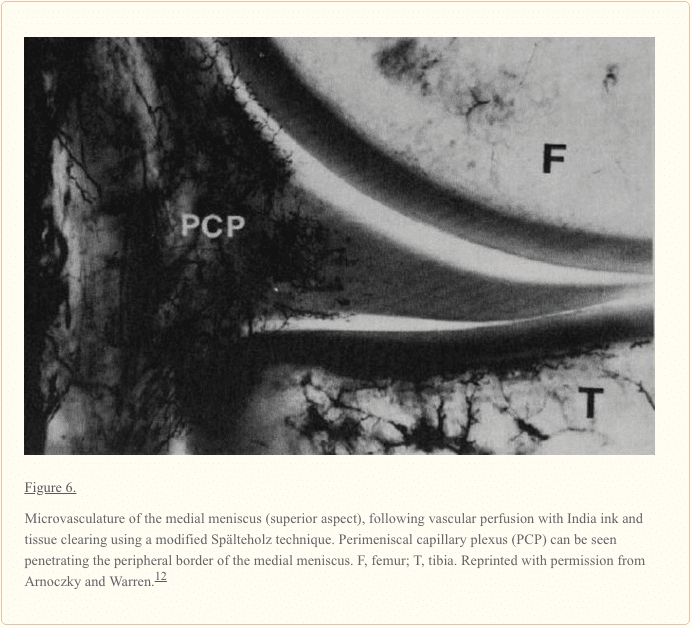
منیسک یک ساختار نسبتا آشکار با یک منبع خون محدود محیطی است. عروق خونی متوسط، جانبی و میانی (که شاخه ای از شریان پاپیتیتال است) عروق کلی را به جنبه های پایین تر و برتر هر منیسک می رساند (شکل 5). 9,12,33-35,148 شریان های خونی متوسط یک شاخه کوچک خلفی است که سوراخ آن رباط پاپیلیتال مچ پا در گوشه پوستی مفصل مفصل تیبوفومورال. یک شبکه مویرگی premeniscal ناشی از شاخه های این شریان ها در داخل بافت های سینوویال و کپسولی زانو در امتداد حاشیه منیسک ایجاد می شود. 10 محیطی٪ تا 30٪ از مرز مدیسال و 10٪ تا 25٪ از منیسک جانبی نسبتا خوبی است که عوارض جانبی دارد که پیامدهای مهمی برای بهبودی منیسک دارد (شکل 6). 12,33,68 عروق انحرافی از شاخهای قدامی و خلفی 33 باقی مانده از هر منیسک (65٪ به 75٪) غذای دریافت شده از مایع سینوویال را از طریق انتشار یا مکانیکی پمپاژ (به عنوان مثال، حرکت مشترک) دریافت می کند. .116,120
پرنده و شیرین منیسک حیوانات و انسان را با استفاده از میکروسکوپ الکترونی و میکروسکوپ نوری بررسی کردند. 23,24 آنها ساختارهای کانال شبیه به عمق سطح منیسک را مشاهده کردند. این کانال ها ممکن است در حمل و نقل مایع در منیسک نقش داشته باشد و ممکن است مواد مغذی را از مایع سینوویال و رگ های خونی به بخش های مغزی از منیسک حمل کند. 23,24 با این حال، مطالعه دقیق تر برای مشخص کردن مکانیزم دقیق که توسط آن حرکت مکانیکی تغذیه به بخش عصبی منیسکی.
کالبدشناسی اعصاب
مفصل زانو توسط شعاع مفصلی خلفی عصب تابیال خلفی و شاخه های پایه عصب های افترتور و فمورال ایجاد می شود. بخش جانبی از کپسول توسط شاخه پروتئین مکرر عصب پرونئال رایج است. این الیاف عصبی به کپسول نفوذ می کنند و از عرضه عروق به قسمت محیطی منیسک و شاخ های قدامی و خلفی که بیشترین الیاف عصبی متمرکز می شوند، دنبال می شود. 52,90 سومین قسمت بیرونی بدن منیسک از دیواره های غلیظ تر است middle third.183,184 در طی شدت خم شدن و گسترش زانو، شاخهای منیسک تحت تأثیر قرار میگیرند و ورودی عصبی در این موقعیتهای شدید بیشتر است. 183,184
گیرنده های مکانیکی درون منیسک به عنوان مبدل عمل می کنند و محرک فیزیکی کشش و فشرده سازی را به یک فشار خاص عصب الکتریکی تبدیل می کنند. مطالعات منیسک انسانی 3 گیرنده مکانیکی از نظر مورفولوژیکی را مشخص کرده است: انتهای روفینی ، پیکرهای پاکیین و اندام های تاندون گلژی. cep گیرنده های مکانیکی نوع I (روفینی) آستانه پایین هستند و به آرامی با تغییرات تغییر شکل و فشار مفصل سازگار می شوند. گیرنده های مکانیکی نوع II (پاچینی) آستانه کم و سریع با تغییرات تنش سازگار می شوند. نوع III (گلژی) گیرنده های مکانیکی آستانه بالا هستند ، که هنگام نزدیک شدن مفصل زانو به دامنه حرکت انتهایی حرکت می کنند و با مهار عصبی عضلانی همراه هستند. این عناصر عصبی با غلظت بیشتری در شاخ های مینیسک ، به ویژه شاخ خلفی پیدا شدند.
اجزای نامتقارن زانو در هماهنگی به عنوان نوعی انتقال بیولوژیکی عمل می کنند که بارهای ران ، استخوان درشت نی ، استخوان کشکک و استخوان ران را می پذیرد ، منتقل می کند. 41 رباط ها به عنوان یک اتصال تطبیقی عمل می کنند ، مینیسک ها نشان دهنده یاتاقان های متحرک هستند. چندین مطالعه گزارش کرده اند که اجزای مختلف درون مفصلی زانو حساس هستند و قادر به تولید سیگنالهای حسی عصبی هستند که به سطح نخاع ، مخچه و بالاتر در سیستم عصبی مرکزی می رسند. اعتقاد بر این است که این سیگنالهای حسی عصبی منجر به درک آگاهانه می شوند و برای عملکرد طبیعی مفصل زانو و حفظ هموستاز بافت مهم هستند. 42

منیسک غضروف است که یکپارچگی ساختاری و عملکردی را به زانو می دهد. Menisci دو پا از بافت فیبر کلاژینو است که گسترش اصطکاک در مفصل زانو را هنگامی که تحت تنش و پیچ خوردگی بین استخوان ران یا تیبیا، و استخوان ران یا فمور است. شناختن آناتومی و بیومکانیک مفصل زانو در درک آسیب های زانو و / یا شرایط ضروری است. دکتر الکس جیمنز DC، CCST Insight
�
عملکرد بیومکانیک
عملکرد بیومکانیکی مینیسک بازتابی از آناتومی ناخالص و فراساختاری و ارتباط آن با ساختارهای داخل مفصلی و خارج مفصلی اطراف است. مینیسک ها بسیاری از عملکردهای مهم بیومکانیکی را انجام می دهند. آنها به انتقال بار ، جذب شوک ، 10,49,94,96,170،51,100,101,109,155،23,24,84,141،102،104,141 ثبات ، 5,15,81,88,115,147،91,172،XNUMX،XNUMX،XNUMX تغذیه ، XNUMX،XNUMX،XNUMX،XNUMX روغنکاری مفصل ، XNUMX-XNUMX،XNUMX و مالکیت کمک می کنند. XNUMX،XNUMX،XNUMX،XNUMX،XNUMX،XNUMX آنها همچنین به منظور کاهش تماس هستند تنشها و افزایش سطح تماس و همزبانی زانو. XNUMX،XNUMX
Kinematics منسکیوم
در مطالعه ای بر روی عملکرد لیگامان، برانتیگان و ووشل گزارش دادند که منیسک داخلی به طور متوسط 2 میلی متر حرکت می کند، در حالی که منیسک جانبی به طور قابل توجهی متحرک تر با تقریباً 10 میلی متر جابجایی قدامی-خلفی در طول خم شدن بود. 25 میلی متر جابجایی قدامی-خلفی را متحمل می شود، در حالی که منیسک جانبی در طول خم شدن 3 میلی متر حرکت می کند.9 در مطالعه ای با استفاده از 37 زانو جسد، تامپسون و همکاران میانگین حرکت داخلی را 5 میلی متر (میانگین شاخ های قدامی و خلفی) گزارش کردند. میانگین حرکت جانبی، 5.1 میلیمتر، در امتداد سطح مفصلی تیبیا (شکل 11.2). یافتههای این مطالعات تفاوت قابلتوجهی را در حرکت سگمنتال بین منیسک داخلی و خارجی تأیید میکند. نسبت منیسک جانبی شاخ قدامی و خلفی کوچکتر است و نشان میدهد که منیسک بیشتر به صورت یک واحد حرکت میکند. همچنین، منیسک داخلی (به طور کلی) کمتر از منیسک جانبی حرکت میکند و حرکت دیفرانسیل شاخ قدامی به خلفی را نشان میدهد. تامپسون و همکاران دریافتند که ناحیه کمترین حرکت منیسک گوشه داخلی خلفی است، جایی که منیسک به دلیل چسبیدن به پلاتوی تیبیا توسط بخش منیسکوتیبیال رباط مایل خلفی محدود می شود، که گزارش شده است که بیشتر مستعد آسیب است. 7 کاهش حرکت شاخ خلفی منیسک داخلی یک مکانیسم بالقوه برای پارگی منیسک است که در نتیجه آن فیبرو غضروف بین کندیل فمور و فلات تیبیا در طول خم شدن کامل به دام میافتد. تفاوت بیشتر بین حرکت شاخ قدامی و خلفی ممکن است منیسک داخلی را در معرض خطر بیشتری برای آسیب قرار دهد.165
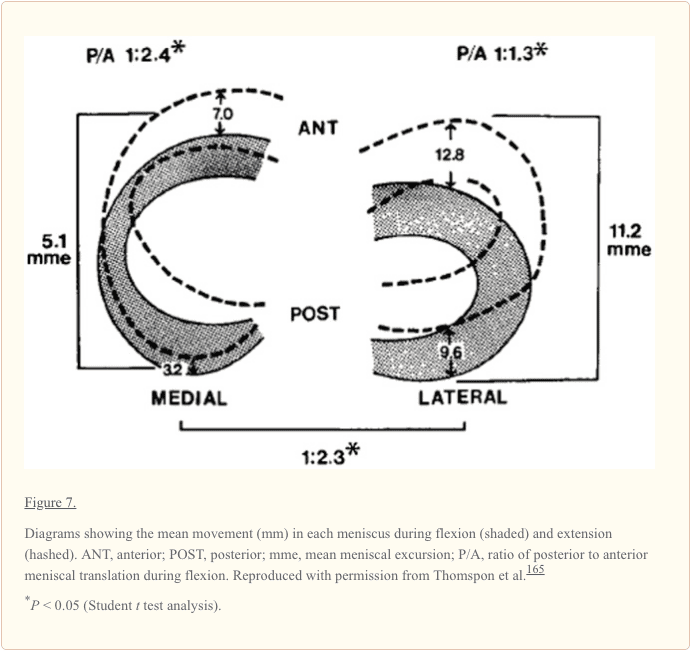
دیفرانسیل شاخ قدامی به حرکت شاخک خلفی اجازه می دهد تا منیسکی به کاهش شعاع با فلکسن، که با شعاع کاهش انحنای فک پایین فمورال خلفی ارتباط دارد. 165 این تغییر شعاع منیسک ها را قادر می سازد تا با سطح مفصل هر دو استخوان ران و تیبیا در طول فلج شدن.
انتقال بار
عملکرد منیسکی به واسطه تغییرات دژنراتیو که همراه آن حذف می شود، به صورت بالینی به دست می آید. Fairbank، افزایش شیوع و تغییرات دژنراتیو پیش بینی شده در سطوح مفصلی در زانوها کاملا منسنکوماتیک را توصیف کرد. 45 از آنجایی که این کار زودهنگام، مطالعات متعدد این یافته ها را تأیید کرده اند و نقش مهم منیسک ها را به عنوان یک ساختار حفاظتی، باربری تقویت کرده اند.
تحمل وزن باعث ایجاد نیروی محوری در سرتاسر زانو می شود که باعث فشرده شدن مینیسک ها و در نتیجه فشارهای هوپی (محیطی) می شود .170 تنش حلقه ای به عنوان نیروهای محوری تولید می شود و در امتداد الیاف کلاژن محیطی مینیسک به تنش های کششی تبدیل می شود (شکل 8). پیوندهای محکم توسط رباط های درونی قدامی و خلفی از اکستروژن مینیسک در محیط تحمل بار جلوگیری می کند. 94 مطالعات Seedhom و Hargreaves گزارش داد که 70٪ بار در محفظه جانبی و 50٪ بار در محفظه داخلی از طریق انتقال می یابد menisci.153 مینیسکها 50٪ از فشار فشاری را از طریق شاخهای خلفی به صورت کششی منتقل می کنند ، و 85٪ آنها را با خم شدن 90 درجه منتقل می کنند. 172 Radin و همکاران نشان دادند که این بارها هنگام سالم بودن مینیسک ها به خوبی توزیع می شوند. 137 با این حال ، حذف منیسک داخلی منجر به کاهش 50 تا 70 درصدی در تماس با کندیل ران ران و افزایش 100 درصدی استرس تماسی می شود. 4,50,91،40،50 کل منیسکتومی جانبی منجر به کاهش 200 تا 300 درصدی در تماس و افزایش استرس تماسی در جز component جانبی به 18,50,76,91 45,85 تا XNUMX of نرمال است. XNUMX،XNUMX،XNUMX،XNUMX این میزان بار در واحد سطح را به طور قابل توجهی افزایش می دهد و ممکن است به تخریب غضروف مفصلی تسریع شده و تحلیل رود. XNUMX،XNUMX
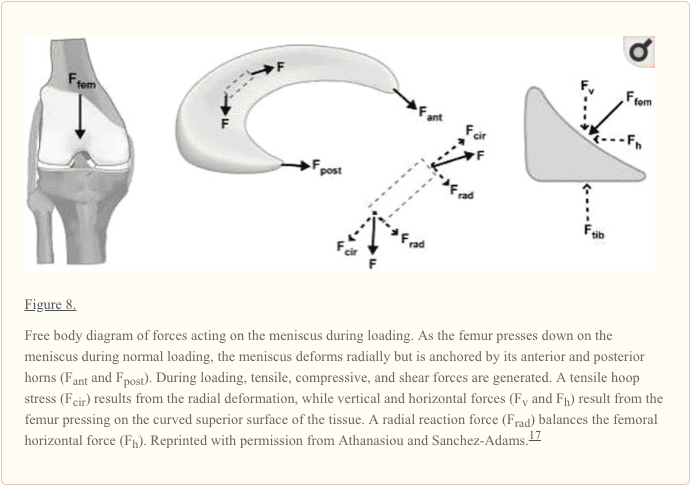
جذب شوک
Menisci نقش مهمی در تضعیف امواج ضربه ای متناوب تولید شده توسط بارگذاری ضربه ای زانو با گام نرمال دارد. 94,96,153 Voloshin و Wosk نشان داد که زانو طبیعی دارای ظرفیت جذب شوک در مورد 20٪ بالاتر از زانو هایی است که تحت منیسککتومی قرار گرفته اند. 170 به عنوان عدم توانایی یک سیستم مشترک برای جذب شوک در توسعه استئوآرتریت منجر شده است، منیسک به نظر می رسد نقش مهمی در حفظ سلامت مفصل زانو داشته باشد. 138
پایداری مشترک
ساختار هندسی منیسک ها نقش مهمی در حفظ سازگاری و ثبات مشترک دارد. ## سطح برتر هر منیسک، سازنده است، که می تواند مفصل موثر بین کاندیداهای فومورو محدب و فلات تیبالی صاف را ایجاد کند. وقتی منیسک دست نخورده است، بارگذاری محوری زانو دارای یک عملکرد ثبات دهنده چند جهته است، محدود کردن حرکت بیش از حد در تمام جهات. 9
مارکولف و همکارانش به تأثیر مینیسکتومی در شلی قدامی - خلفی و چرخشی زانو پرداخته اند. مینیسکتومی داخلی در زانوی دست نخورده ACL تأثیر کمی در حرکت قدامی - خلفی دارد ، اما در زانوی کمبود ACL ، منجر به افزایش ترجمه تیبیای قدامی - خلفی تا 58٪ در 90 درجه خم شدن می شود. 109 Shoemaker and Markolf نشان داد که شاخ خلفی مینیسک داخلی مهمترین ساختار مقاوم در برابر نیروی قدامی استخوان درشتن در زانوی کمبود ACL است .155 آلن و همکاران نشان دادند که نیروی حاصل در مینیسک داخلی زانوی کمبود ACL 52٪ در امتداد کامل و با 197٪ در 60 درجه خم شدن تحت بار قدامی استخوان درشت نی 134-N. 7 تغییرات بزرگ در سینماتیک به دلیل منیسکتومی داخلی در زانوی کمبود ACL نقش مهم مینیسک داخلی را در پایداری زانو تأیید می کند. اخیراً ، Musahl و همکاران گزارش داده اند که منیسک جانبی در مانور چرخش محوری در ترجمه قدامی تیبیا نقش دارد .123
تغذیه و روانکاری مشترک
Menisci همچنین ممکن است در تغذیه و روانکاری مفصل زانو نقش داشته باشد. مکانیسم این روانکاری هنوز معلوم نیست منیسک ممکن است سیال سینوویال را در غضروف مفصلی فشرده کند، که نیروهای اصطکاک را در طول وزن کم می کند. 13
یک سیستم میکرو کانال ها در داخل منیسک واقع در نزدیکی رگ های خونی وجود دارد که با حفره سینوویال ارتباط دارد؛ این ممکن است حمل و نقل مایع برای تغذیه و روانکاری مشترک. 23,24
خودآزاری
درک حرکت مشترک و موقعیت (proprioception) توسط mechanoreceptors متصل است که تغییر شکل مکانیکی را به سیگنال های عصبی الکتریکی انتقال می دهد. Mechanoreceptors در شاخهای قدامی و خلفی منیسک شناسایی شده است. *** Mechanoreceptors سازگاری سریع مانند Corpuscles پچینین، به نظر می رسد که احساس حرکت حرکتی و گیرنده های تطبیق آهسته مانند Endings Ruffini و تاندون Golgi اعتقاد بر این است که اندام ها احساس می کنند که موقعیت مشترک دارند. 140 شناسایی این عناصر عصبی (که عمدتا در سومین وسطی و بیرونی منیسک قرار دارد) نشان می دهد که منیسک ها قادر به شناسایی اطلاعات مفصل در مفصل زانو هستند، نقش مهمی در مکانیسم بازخورد حسی زانو. 61,88,90,158,169
بلوغ و پیری منیسک
میکروآناتومی منیسک پیچیده است و قطعا تغییرات پیری را نشان می دهد. با افزایش سن، منیسک سخت تر می شود، الاستیک را از بین می برد و زرد می شود. 78,95 با میکروسکوپیک، عادت های سلولی با فضاهای خالی و افزایش بافت فیبری در مقایسه با بافت الاستیک به وجود می آید. 74 این مناطق کیستیک می تواند یک پارگی ایجاد کند ، و با نیروی کششی توسط کاندیل فمورال، لایه های سطحی منیسک ممکن است از لایه ی عمیق در رابط تغییر دژنراتیو کیستی برداشته شود و یک پاره ی افقی را ایجاد کند. برش بین این لایه ها ممکن است موجب درد شود. منیسک پاره شده ممکن است به طور مستقیم آسیب برسد غضروف مفصلی. 74,95
غوش و تیلور دریافتند که غلظت کلاژن از بدو تولد به 30 سال افزایش یافته و تا 80 سالگی ثابت مانده است و پس از آن کاهش یافت. 58 پروتئین های ماتریس غیر کلاژن بیشترین تغییرات عمیق را نشان دادند ، از 21.9/1.0٪ 8.1٪ (وزن خشک) کاهش می یابد در نوزادان بین سنین 0.8 تا 30 سال به 70/80 70. 11.6. رسیده است. 1.3 بعد از 131 سالگی ، سطح پروتئین ماتریس غیر کلاژن به XNUMX XNUMX XNUMX/XNUMX increased افزایش یافت. پیترز و اسمیلی با افزایش سن افزایش هگزوزامین و اسید اورونیک را مشاهده کردند
McNicol و Roughley تنوع پروتئگلیکان های منسک را در ageing113 بررسی کردند؛ تفاوت های کوچک در میزان استخراج و اندازه هیدرودینامیکی مشاهده شد. نسبت سنتز کراتین نسبت به کلردروتین-6-سولفات با افزایش سن افزایش می یابد. 146
پترسن و تیلمن از نظر ایمونوهیستوشیمی با مشاهده تمایز رگهای خونی و لنفاوی در 22 جسد انسانی ، مینیسک های انسانی (از 80 هفته حاملگی تا 20 سال) را بررسی کردند. در زمان تولد ، تقریباً کل مینیسک واسکولار شد. در سال دوم زندگی ، یک ناحیه عروقی در محیط داخلی ایجاد شد. در دهه دوم ، رگهای خونی در یک سوم محیطی وجود داشتند. پس از 50 سال سن ، فقط یک چهارم محیطی قاعده مینیسک واسکولار شد. بافت پیوندی متراکم محل قرارگیری ، واسکولار شد اما غضروف فیبروکارتال داخل آن نیست. رگهای خونی در تمام مناطق با لنفاوی همراه بودند
Arnoczky پیشنهاد کرد که وزن بدن و حرکت مفصل زانو ممکن است رگهای خونی را در جنبه های داخلی و میانی منیسک از بین ببرند. 9 تغذیه بافت منیسک از طریق پرفیوژن از عروق خونی و از طریق انتشار سیال سینوویال اتفاق می افتد. نیاز به تغذیه از طریق انتشار، بارگیری متناوب و آزاد شدن روی سطوح مفصلی است که تحت تأثیر وزن بدن و نیروهای عضلانی است. 130 مکانیسم با تغذیه غضروف مفصلی قابل مقایسه است. 22
تصویربرداری رزونانس مغناطیسی منیسک
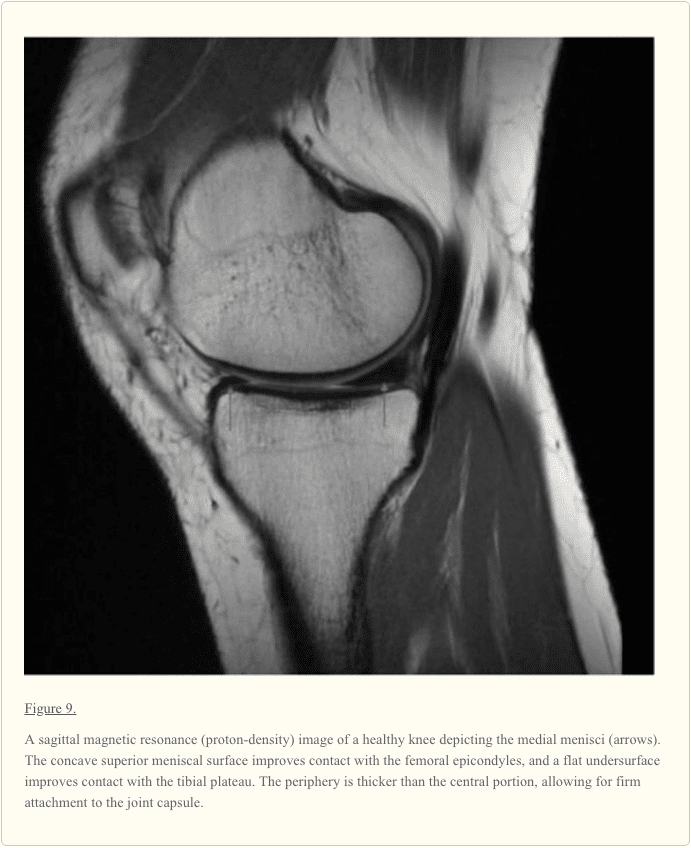
تصویربرداری رزونانس مغناطیسی (MRI) ابزار تشخیصی غیرمخرب است که در ارزیابی، تشخیص و نظارت بر منیسک استفاده می شود. به علت کنتراست بافت نرم نسبتا بالا، MRI به عنوان روش مرسوم تصویربرداری به طور گسترده پذیرفته شده است.
در MRI متقاطع، منیسک طبیعی به عنوان یک ساختار مثلثی با فرمول پایین (تاریک) یکنواخت (شکل 9) ظاهر می شود. پارگی منسکی با وجود افزایش سیگنال داخلی که به سطح این ساختار گسترش می یابد شناسایی می شود.
مطالعات متعددی، ابزار بالینی MRI را برای اشک منصری ارزیابی کرده اند. به طور کلی، MRI برای اشک های منیسک بسیار حساس و خاص است. حساسیت MRI در تشخیص اشکاهای منیسک از 70٪ به 98٪ و مشخصه از 74٪ تا 98٪ است. 48,62,105,107,117 MRI بیماران 1014 قبل از بررسی آرتروسکوپی، دقت 89٪ برای آسیب شناسی منیسک مدیا و 88٪ برای منیسک جانبی.48 یک متاآنالیز از بیماران 2000 با MRI و معاینه آرتروسکوپی حساسیت 88٪ و دقت 94٪ برای اشک منیسک X.NUMX
اختلافاتی بین تشخیص MRI و آسیب شناسی مشخص شده در حین بررسی آرتروسکوپی وجود دارد. Justice و Quinn اختلافاتی را در تشخیص 66 مورد از 561 بیمار (12٪) گزارش کردند. 86 در مطالعه ای بر روی 92 بیمار ، اختلاف بین MRI و در 22 مورد از 349 مورد (6٪) تشخیص های آرتروسکوپی مشاهده شد .106 میلر یک مطالعه آینده نگر تک کور را با مقایسه معاینات بالینی و MRI در 57 معاینه زانو انجام داد. 117 وی تفاوت معنی داری در حساسیت بین معاینه بالینی و MRI (80.7) مشاهده نکرد. به ترتیب و 73.7٪). Shepard و همکاران دقت MRI را در شناسایی ضایعات بالینی قابل توجه شاخ قدامی مینیسک در 947 زانو متوالی MRI154 ارزیابی کردند و میزان مثبت کاذب 74٪ را یافتند. افزایش شدت سیگنال در شاخ قدامی لزوماً نشان دهنده ضایعه بالینی قابل توجه نیست
نتیجه گیری
منیسک مفصل زانو، قاعده های شیب دار فیبروکولاژیک است که باعث افزایش ثبات در مفصل فموروتبیل می شود، بار محوری را بارگذاری می کند، شوک جذب می کند و روانکاری به مفصل زانو را فراهم می کند. آسیب های منیسکی به عنوان یک علت عوارض اسکلتی عضلانی شناخته شده است. حفاظت از منیسکی به شدت وابسته به حفظ ترکیب و سازمان متمایز آن است.
سپاسگزاریها
Ncbi.nlm.nih.gov/pmc/articles/PMC3435920/
پانویسها و منابع
Ncbi.nlm.nih.gov/pmc/articles/PMC3435920/
در نتیجه ، زانو بزرگترین و پیچیده ترین اتصال بدن انسان است. با این حال ، از آنجا که زانو معمولاً در اثر آسیب و یا شرایطی ممکن است آسیب ببیند ، درک آناتومی مفصل زانو برای اینکه بیماران بتوانند درمان مناسب داشته باشند ضروری است. scope دامنه اطلاعات ما محدود به کایروپراکتیک و مسائل مربوط به سلامت ستون فقرات. برای بحث در مورد موضوع ، لطفاً از دکتر جیمنز سوال کنید یا با ما تماس بگیرید915-850-0900.
دکتر الکس جیمنز سرپرستی می کند

بحث موضوعی اضافی: کاهش درد زانو بدون جراحی
زانو درد یک علامت شناخته شده است که می تواند به دلیل آسیب دیدگی های مختلف زانو و / یا شرایط ، از جمله رخ دهدآسیب های ورزشی. زانو یکی از پیچیده ترین اتصالات بدن انسان است که از تقاطع چهار استخوان، چهار رباط، تاندون های مختلف، دو منیسکی و غضروف ساخته شده است. به گفته آکادمی پزشکان خانواده آمریکایی، شایع ترین علل درد زانو شامل سوزش پاتلا لار، تاندونیت پاتلا و یا زانو بلوز و بیماری Osgood-Schlatter می باشد. گرچه درد زانو بیشتر در افراد بالای 60 رخ می دهد، درد زانو نیز در کودکان و نوجوانان رخ می دهد. بعد از روش های RICE، درد زانو را می توان در خانه درمان کرد، با این حال، آسیب های شدید زانو ممکن است نیاز به مراقبت فوری پزشکی، از جمله مراقبت از مراقبت از ماساژ.

EXTRA EXTRA | موضوع مهم: El Paso، TX Chiropractor Recommended
جای خالی
منابع
بستن آکاردئون
اطلاعات اینجا در "علم پایه ساختار، ترکیب، و عملکرد Menisci زانو انسان"در نظر گرفته شده است که جایگزین رابطه یک به یک با یک متخصص مراقبت های بهداشتی واجد شرایط یا پزشک دارای مجوز نیست و توصیه پزشکی نیست. ما شما را تشویق می کنیم که تصمیمات مراقبت های بهداشتی را بر اساس تحقیقات و مشارکت خود با یک متخصص مراقبت های بهداشتی واجد شرایط اتخاذ کنید.
محدوده اطلاعاتی ما محدود به کایروپراکتیک، اسکلتی عضلانی، داروهای فیزیکی، سلامتی، کمک کننده به علت اختلالات احشایی در ارائه های بالینی، پویایی بالینی رفلکس سوماتوویسرال مرتبط، کمپلکس های سابلوکساسیون، مسائل حساس سلامتی، و/یا مقالات، موضوعات و بحث های پزشکی کاربردی.
ارائه و ارائه می کنیم همکاری بالینی با متخصصین رشته های مختلف هر متخصص بر اساس حوزه فعالیت حرفه ای و صلاحیت مجوز آنها اداره می شود. ما از پروتکل های عملکردی سلامت و تندرستی برای درمان و حمایت از مراقبت از آسیب ها یا اختلالات سیستم اسکلتی عضلانی استفاده می کنیم.
ویدیوها، پستها، موضوعات، موضوعات و بینشهای ما، موضوعات، مسائل و موضوعات بالینی را پوشش میدهد که به طور مستقیم یا غیرمستقیم به حوزه عمل بالینی ما مربوط میشود و به طور مستقیم یا غیرمستقیم از آن پشتیبانی میکند.*
دفتر ما به طور منطقی تلاش کرده است تا استنادات حمایتی ارائه دهد و مطالعه تحقیقاتی یا مطالعات مرتبط با پست های ما را شناسایی کرده است. ما کپی از مطالعات تحقیقاتی پشتیبانی را که در صورت درخواست در دسترس هیئت های نظارت و عموم است ، ارائه می دهیم.
ما می فهمیم که مواردی را پوشش می دهیم که نیاز به توضیح اضافی در مورد چگونگی کمک به آن در یک برنامه مراقبت خاص یا پروتکل درمانی دارند. بنابراین ، برای بحث بیشتر در مورد موضوع فوق ، لطفاً آزادانه س .ال کنید دکتر الکس جیمنز، دی سی, و یا با ما تماس بگیرید در 915-850-0900.
ما برای کمک به شما و خانواده شما اینجا هستیم.
نعمت
دکتر الکس جیمنز DC ، MSACP, RN*, CCST, IFMCP*, CIFM*, ATN*
ایمیل شما: coach@elpasofunctionalmedicine.com
دارای مجوز به عنوان دکتر کایروپراکتیک (DC) در وابسته به تکزاس & نیومکزیکو*
مجوز تگزاس دی سی شماره TX5807, نیومکزیکو دی سی مجوز # NM-DC2182
دارای مجوز به عنوان پرستار ثبت شده (RN*) in فلوریدا
مجوز RN مجوز فلوریدا # RN9617241 (شماره کنترل 3558029)
وضعیت فشرده: مجوز چند ایالتی: مجاز به تمرین در کشورهای 40*
دکتر الکس جیمنز DC، MSACP، RN* CIFM*، IFMCP*، ATN*، CCST
کارت ویزیت دیجیتال من





