تیم پزشکی عملکردی اپی ژنتیک کلینیک برگشت. مطالعه تغییرات ارثی در بیان ژن (ژنهای فعال در مقابل ژنهای غیرفعال) شامل تغییراتی در توالی DNA نیست، تغییر در فنوتیپ بدون تغییر در ژنوتیپ، که بر نحوه خواندن ژنها توسط سلولها تأثیر میگذارد. تغییر اپی ژنتیک یک اتفاق طبیعی و منظم است که می تواند تحت تأثیر عوامل متعددی قرار گیرد: سن، محیط، شیوه زندگی و وضعیت بیماری. تغییرات اپی ژنتیکی معمولاً می تواند به صورت تمایز نهایی سلول ها به سلول های پوست، سلول های کبد، سلول های مغز و غیره ظاهر شود. و تغییرات اپی ژنتیکی می تواند اثرات مخرب تری داشته باشد که می تواند منجر به بیماری شود.
تحقیقات جدید و در حال انجام به طور مداوم نقش اپی ژنتیک را در انواع اختلالات انسانی و بیماری های کشنده کشف می کند. علائم اپی ژنتیک در بزرگسالی پایدارتر است. با این حال، هنوز تصور می شود که آنها با انتخاب سبک زندگی و تأثیر محیطی پویا و قابل تغییر هستند. آشکار می شود که اثرات اپی ژنتیک نه تنها در رحم بلکه در طول زندگی انسان رخ می دهد. کشف دیگر این است که تغییرات اپی ژنتیکی را می توان معکوس کرد. نمونه های متعددی از اپی ژنتیک نشان می دهد که چگونه انتخاب های مختلف سبک زندگی و قرار گرفتن در معرض محیطی می تواند علائم روی DNA را تغییر دهد و در تعیین نتایج سلامتی نقش داشته باشد.
چگونه تغذیه اپی ژنتیکی و شخصی شده به سلامتی بهینه کمک می کند؟
اکثر ما درباره غذاهای ناسالم می دانیم که چگونه بدن ما را تحت تاثیر قرار می دهد. آنها
متابولیسم را کند کنید
وزن اضافه کنید
شریان های سخت و پیچیده و غیره
اما اکنون مواد غذایی و عناصر غذای وجود دارد که می تواند به ما کمک کند و از جایی که ممکن است ما فکر نکنیم، و این DNA ما است.
Nutriepigenomics بررسی ارتباط بین رژیم غذایی و biomarkers که می تواند متصل شده و یا از DNA ما حذف شده است. این ژنهای ما را روشن یا خاموش می کند.
مطالعات جدید نشان می دهد که مشخص است غذاها یا مکمل ها می تواند بیان ژن های ما را تنظیم کند که می تواند سلامت ما را تحت تاثیر قرار دهد.
ژنومیک تغذیه انقلابی در شیوه های تغذیه بالینی و بهداشت عمومی است:
رژیم غذایی، ورزش، و قرار گرفتن در معرض قرار گرفتن در معرض محیط زیست، همه عناصری هستند که نقش مهمی در تعویض ژن ها از طریق اپی ژنتیک دارند. تنظیم عوامل سبک زندگی می تواند توانایی کاهش بیماری را کنترل و تاثیر مثبتی بر سلامت ما داشته باشد.
متخصصین سلامت از همه چیز شروع به ترکیب epigenetics به عمل خود را با هدف ارائه برنامه های تخصصی تر و فردی تر درمان.
Kristy Hall ، MS ، RNCP ، ROHP ، یک هیئت مدیره متخصص تغذیه عملکرد و گفت: "اطلاعات لایه بندی مانند رژیم غذایی ، سبک زندگی ، عوامل محیطی ، سابقه خانوادگی ، علائم و تشخیص ها بهمراه اپی ژنتیک می تواند به هدایت کسی به وضعیت سلامت بهینه کمک کند." بنیانگذار Living Well Nutrition که از آزمایش اپی ژنتیک ، مشاوره تغذیه و یک رویکرد چند وجهی برای تأمین بهتر مشتریان خود استفاده می کند.
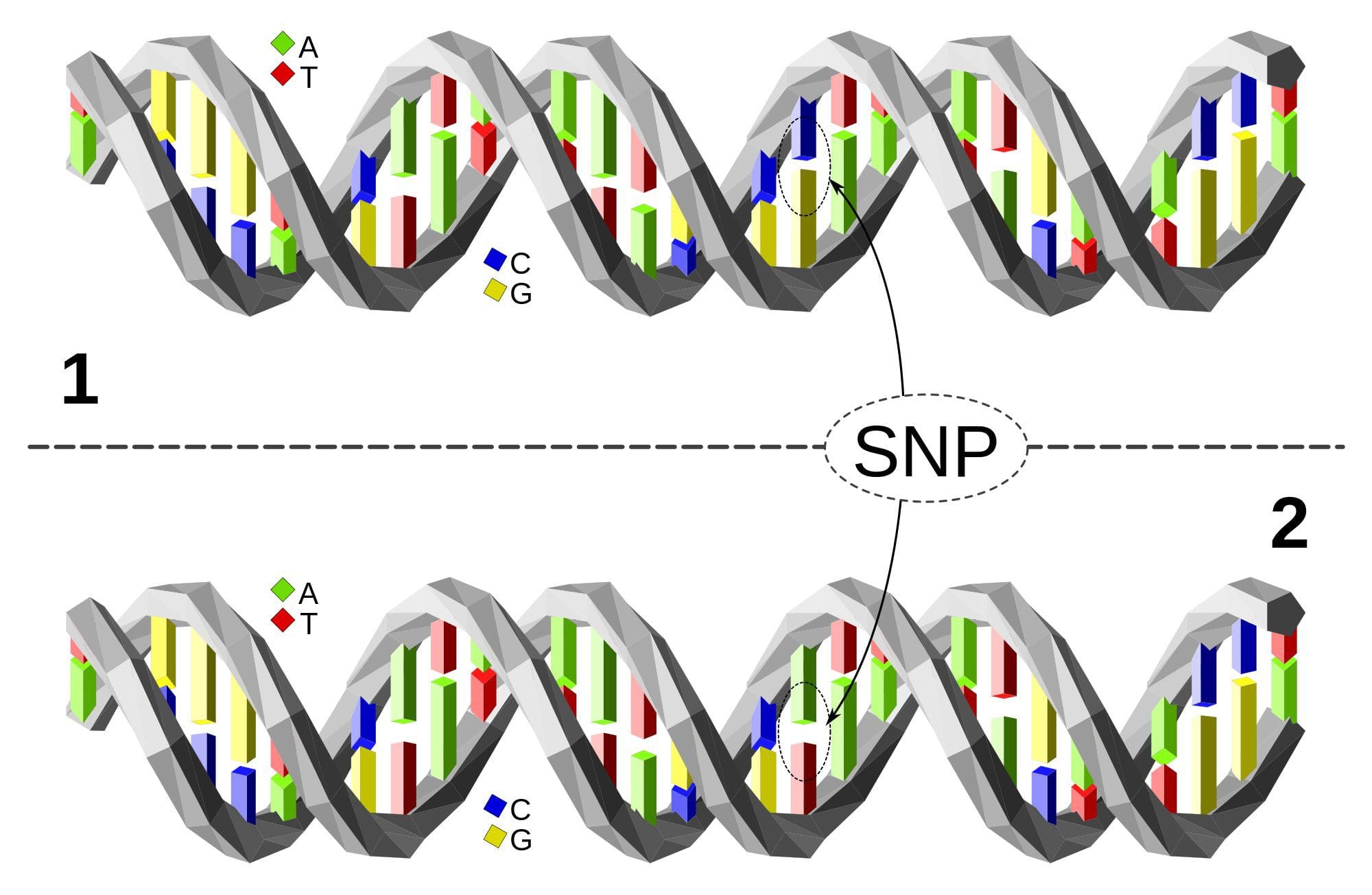
مولکول DNA فوقانی از مولکول DNA پایین تر در یک محل واحد جفت پایه (پلی مورفیسم C / A) متفاوت است
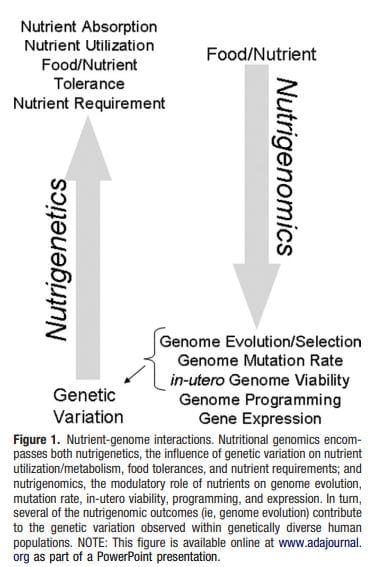
ژنتیک تغذیه ای یا nutrigenetics شامل شناسایی، طبقه بندی و مشخصه از تغییرات ژنتیکی انسان است که متابولیسم مواد مغذی / مصرف و تحمل مواد غذایی Fig1 را تغییر می دهد.
IOM Nutrigenomics و فراتر از آن: آگاهی از آینده. واشنگتن دی سی: مطبوعات ملی آکادمی؛ 2007
کاربرد: ژنتیک و اپی ژنتیک
برای مثال، مواد مغذی دارو، اثر سازنده قدرت بیان و ثبات ژنوم هستند و این تعاملات ژنتیکی مواد مغذی را می توان برای جلوگیری از بیماری ها بهینه سازی کرد.
تغذیه فردی
وعده مهندسی تغذیه برای سلامتی مطلوب از طریق رژیم غذایی هنوز در حال انجام است، اما مردم انتظارات مثبتی را در اختیار دارند، همانطور که با استفاده از مکمل های غذایی حاكی است.
تحقیقات علمی نشان می دهد که مواد مغذی در غذاهای مختلف و مکمل های غذایی که ما می خوریم ممکن است قادر به تغییر یا تغییر مقادیر سرطانی باشند. این شواهد را می توان در انتخاب شیوه زندگی بهتر استفاده کرد.
زغال اخته فوق العاده سرشار از آنتی اکسیدان است و تصور می شود که این "سوپرفود" می تواند از نظر ژنتیکی باعث کاهش آسیب DNA شود ، در نتیجه از انسان در برابر سرطان و حتی حتی پیری آهسته محافظت می کند. آب زغال اخته و ویتامین C به عنوان مهارکننده های بالقوه متیلاسیون ژن MTHFR و ژن DNMT1 در انسان نشان داده شده است.
Kim، M.، Na، H.، Kasai، H.، Kawai، K.، Li، Y.-S.، & Yang، M. (2017). مقایسه زغال اخته (Vaccinium spp.) و ویتامین C از طریق اثرات آنتی اکسیدانی و اپی ژنتیکی در انسان. مجله پیشگیری از سرطان ، 22 (3) ، 174 181.
یادگیری در مورد آنچه که ما می خوریم و آنچه که به بدن ما می انجامد، به ویژه تاثیرات بالقوه اپی ژنتیک، تنها یک قدم به سلامت مطلوب است.
شيوع افزايش چاقي و ارتباطات مرتبط با آن يكي از مشكلات مهم بهداشت عمومي است. اگرچه عوامل ژنتیکی در تعیین حساسیت فرد به افزایش وزن و چاقی نقش مهمی دارند، اما انواع ژنتیکی شناسایی تنها بخشی از تغییرات را توضیح می دهند. این به نفع رو به رشدی در درک نقش بالقوه اپی ژنتیک به عنوان واسطه کننده تعاملات ژن-محیط زیست در زمینه توسعه چاقی و ارتباطات مرتبط با آن منجر شده است. شواهد اولیه در حمایت از نقش epigenetics در چاقی و نوع diabetes mellitus 2 (T2DM) عمدتا توسط مطالعات حیوانی ارائه شده است که تغییرات اپی ژنتیکی در بافت های مهم کلیدی متابولیک را پس از تغذیه با چربی زیاد و تفاوت های اپی ژنتیکی بین حیوانات نابینا و چاق و توسط مطالعات انسانی که نشان دهنده تغییرات اپی ژنتیکی در چاقی و ژنهای کاندیدای T2DM در افراد چاق / دیابتی بود. به تازگی، پیشرفت های روش های اپی ژنتیک و کاهش هزینه های مطالعات ارتباط گسترده ایپیمنوم (EWAS) منجر به گسترش سریع مطالعات در جمعیت های انسانی شده است. این مطالعات همچنین تفاوت های اپيگنتیک بین بزرگسالان چاق / T2DM و کنترل های سالم و تغییرات اپی ژنتیکی در ارتباط با تغذیه، کاهش وزن و مداخلات ورزشی گزارش شده است. همچنین شواهد در حال افزایش از مطالعات انسان و حیوانات وجود دارد که ارتباط بین اثرات تغذیه ای پریناتال و خطر ابتلا به چاقی و T2DM ممکن است با تغییرات اپيگنتیک در فرزندان موثر باشد. هدف از این بررسی خلاصه آخرین تحولات در این زمینه به سرعت در حال حرکت است، با توجه خاص به EWAS انسانی و مطالعات بررسی تاثیر عوامل تغذیه ای و سبک زندگی (هر دو قبل و بعد از تولد) بر epigenome و ارتباط آنها با متابولیسم نتایج سلامت مشکلات در تشخیص نتیجه از علیت در این مطالعات و نقش حیاتی مدل های حیوانات برای آزمایش روابط علی و ارائه بینش به مکانیزم های زمینه ای نیز مورد توجه قرار گرفته است. به طور خلاصه، مساحت اپی ژنتیک و سلامت متابولیسم، در فواصل کوتاهی، پیشرفت های سریع را تجربه کرده است. در حالی که نتایج تا به امروز امیدوار کننده است، مطالعات در حال انجام است، و دهه آینده وعده می شود که زمان تحقیقات مولد در تعامل پیچیده بین ژنوم، epigenome و محیط زیست به عنوان آنها مربوط به بیماری متابولیکی است.
کلید واژه ها: Epigenetics، متیلاسیون DNA، چاقی، نوع 2 دیابت، برنامه ریزی توسعه
معرفی
چاقی یک بیماری پیچیده و چندوجهی است و درک بهتر مکانیسم های درونی متابولیسم زندگی، محیط زیست و ژنتیک برای ایجاد راهکارهای موثر برای پیشگیری و درمان [1] ضروری است.
در جامعهای که مواد غذایی پرانرژی فراوان و نیاز به فعالیت بدنی کم است، تنوع زیادی در استعداد ابتلا به چاقی و مشکلات سلامت متابولیک وجود دارد. تخمینها از نقش وراثت در این تنوع در محدوده 40 تا 70 درصد است و در حالی که مطالعات ارتباط گسترده ژنومی (GWAS) تعدادی مکان ژنتیکی مرتبط با خطر چاقی را شناسایی کردهاند، تنها 100 نوع ژنتیکی رایج هستند. چند درصد از واریانس چاقی را تشکیل می دهند [2، 3]. برآوردهای کل ژنوم بالاتر است و حدود 20 درصد از تغییرات را شامل می شود [3]. با این حال، بخش بزرگی از وراثت ناشناخته باقی مانده است.
اخیراً توجه به بررسی نقش تغییرات اپی ژنتیکی در اتیولوژی چاقی معطوف شده است. استدلال شده است که اپی ژنوم ممکن است نشان دهنده پیوند مکانیکی بین گونه های ژنتیکی و عوامل محیطی در تعیین خطر چاقی باشد و می تواند به توضیح "وراثت ناپدید شده" کمک کند. اولین مطالعات اپی ژنتیکی انسانی کوچک بودند و فقط تعداد محدودی مکان را بررسی کردند. در حالی که این به طور کلی منجر به تکرارپذیری ضعیف می شود، برخی از این یافته های اولیه، به عنوان مثال، رابطه بین متیلاسیون PGC1A و دیابت نوع 2 (T2DM) [4] و دیگران همانطور که در ون دایک و همکاران بحث شده است. [5]، در مطالعات بعدی تکرار شده است. پیشرفتهای اخیر و افزایش قیمتدهی فناوریهای با توان بالا، اکنون امکان انجام مطالعات ارتباط گسترده اپی ژنوم (EWAS) و ادغام لایههای مختلف اطلاعات ژنومی را برای کشف تعاملات پیچیده بین ژنوتیپ، اپی ژنوم، رونوشت و محیط فراهم میکند. 6]. این مطالعات هنوز در مراحل ابتدایی خود هستند، اما نتایج تا کنون نشان داده است که به توضیح تنوع در حساسیت به چاقی کمک می کند.
شواهد فزاینده ای وجود دارد که نشان می دهد چاقی ریشه ذهنی دارد ، زیرا قرار گرفتن در معرض یک منبع غذایی بهینه بهینه قبل از تولد یا در اوایل نوزادی با افزایش خطر چاقی و بیماری های متابولیکی در زندگی بعدی مرتبط است [10-13]. در ابتدا ، مطالعات حیوانی نشان داد که طیف وسیعی از مواجهه های تغذیه ای در اوایل زندگی ، به ویژه مواردی که در اوایل بارداری تجربه کرده اند ، می توانند تغییرات اپی ژنتیکی را در بافتهای متابولیکی اصلی فرزندان ایجاد کنند که پس از تولد نیز ادامه داشته و منجر به تغییرات دائمی در عملکرد ژن می شوند [13 17]. شواهدی در دست اثبات وجود مکانیسم مشابه در انسان است. این امر منجر به جستجوی علائم اپی ژنتیکی موجود در اوایل زندگی شده است که خطر بعدی بیماری متابولیک را پیش بینی می کند و مطالعاتی برای تعیین اینکه آیا برنامه ریزی اپی ژنتیکی بیماری متابولیک می تواند در زندگی بعدی جلوگیری یا معکوس شود ، تعیین می کند.
این بررسی به روز رسانی از بررسی قبلی سیستماتیک مطالعات در مورد اپی ژنتیک و چاقی در انسان می پردازد [5]. بررسی قبلی ما نتایج امیدوار کننده مطالعات اولیه را نشان داد، از جمله اولین علائم بالقوه اپی ژنتیک برای چاقی است که می تواند در هنگام تولد تشخیص داده شود (به عنوان مثال، RXRA) [18]. با این حال، آن را نیز نشان داد که بازدهی محدود از یافته ها و عدم تحقیقات طولی مقیاس بزرگتر. بررسی کنونی بر پیشرفت های اخیر در این زمینه به سرعت در حال حرکت و به ویژه در EWAS انسانی و مطالعاتی پیرامون تأثیر عوامل (پیش و پس از تولد) تغذیه و شیوه زندگی بر اپژئوم و نقش در حال ظهور اپی ژنتیک در آسیب شناسی چاقی متمرکز است. . ما همچنین به مشکلات مربوط به شناسایی علیت در این مطالعات و اهمیت مدل های حیوانی در ارائه بینش به مکانیسم ها پرداخته ایم.
مرور
تغییرات قارچی در مدل های حیوانی چاقی
مدلهای حیوانی فرصتهای منحصر به فردی را برای مطالعات بسیار کنترلشده فراهم میکنند که بینش مکانیکی در مورد نقش علائم اپی ژنتیک خاص، هم به عنوان شاخصهای وضعیت متابولیک فعلی و هم بهعنوان پیشبینیکننده خطر آتی چاقی و بیماریهای متابولیک ارائه میکنند. یکی از جنبههای مهم مطالعات حیوانی این است که آنها امکان ارزیابی تغییرات اپی ژنتیکی در بافتهای هدف، از جمله کبد و هیپوتالاموس را فراهم میکنند، که در انسان بسیار دشوارتر است. علاوه بر این، توانایی برداشت مقادیر زیادی از بافت تازه، ارزیابی چندین علامت کروماتین و همچنین متیلاسیون DNA را ممکن میسازد. برخی از این تغییرات اپی ژنتیکی به تنهایی یا به صورت ترکیبی ممکن است به برنامه ریزی محیطی پاسخ دهند. در مدلهای حیوانی، میتوان چندین نسل از فرزندان را مطالعه کرد و بنابراین تمایز بین انتقال بین نسلی و بین نسلی خطر چاقی را با واسطه حافظه اپی ژنتیکی وضعیت تغذیه والدین، که در مطالعات انسانی به راحتی قابل تشخیص نیست، امکانپذیر کرد. ما از اصطلاح اول برای انتقال میوز خطر در غیاب مواجهه مداوم استفاده می کنیم در حالی که دومی در درجه اول مستلزم انتقال مستقیم خطر از طریق برنامه ریزی مجدد متابولیک جنین یا گامت است.
مطالعات حیوانی نقش مهمی در درک فعلی ما از نقش اپی ژنتیک در ریشه رشد چاقی و T2DM داشته است. افزایش و کاهش تغذیه مادر در دوران بارداری با افزایش رسوب چربی در فرزندان بیشتر گونه های پستانداران مورد مطالعه تاکنون همراه بوده است (بررسی شده در [11 ، 13-15 ، 19]). تغذیه مادران در دوران بارداری نه تنها پتانسیل تأثیر مستقیم بر جنین را دارد ، بلکه ممکن است مستقیماً بر تخمک های در حال رشد جنین های ماده و سلول های زایای اولیه جنین های نر نیز تأثیر بگذارد و بنابراین می تواند بر فرزندان خارج از بهار و بزرگ تأثیر بگذارد. از این رو ، برای ایجاد تمایز بین مکانیسم های انتقال بین نسلی و انتقال نسلی مادر ، داده های چند نسلی معمولاً مورد نیاز است.
جدول 1 خلاصه انواع مدل های حیوانی است که برای نشان دادن تغییرات متابولیکی و اپی ژنتیکی در فرزندان همراه با سطح والدین تغذیه استفاده شده است. همچنین حاوی اطلاعات مربوط به مطالعات برای شناسایی علامت های اپی ژنتیک تغییر یافته در افراد بالغ که به چالش های تغذیه مستقیم می اندیشند. جدول توسط نوع انتقال خطر پیشنهاد شده است.
(I) تغييرات اپيگنتيک در پسران همراه با تغذيه مادر در حين بارداري
مکمل های غذایی مادر، سوء تغذیه و تغذیه بیش از حد در دوران بارداری می تواند رسوب چربی و هموستاز انرژی را در فرزندان تغییر دهد [11، 13، 15، 19]. با این اثرات در فرزندان، تغییرات در متیلاسیون DNA، تغییرات پس از ترجمه هیستون، و بیان ژن برای چندین ژن هدف، به ویژه ژن های تنظیم کننده متابولیسم اسیدهای چرب و سیگنال دهی انسولین مرتبط است [16، 17، 20-30]. تنوع مدلهای حیوانی مورد استفاده در این مطالعات و مسیرهای متابولیکی رایج تحت تأثیر قرار گرفته، نشاندهنده یک پاسخ تطبیقی حفظشده تکاملی است که با اصلاح اپی ژنتیکی انجام میشود. با این حال، تعداد کمی از ژنهای شناسایی شده خاص و تغییرات اپی ژنتیکی در مطالعات مرتبط تایید شدهاند، و تحقیقات گسترده در مقیاس ژنوم معمولاً اعمال نشدهاند. یک مانع عمده برای مقایسه این مطالعات، پنجرههای ذهنی مختلف است که در معرض چالشهای تغذیهای قرار میگیرند، که ممکن است نتایج بسیار متفاوتی را ایجاد کند. اثبات اینکه تغییرات اپی ژنتیکی به جای اینکه با تغییرات فنوتیپی فرزندان مرتبط باشد، علت و معلول هستند نیز مورد نیاز است. این امر مستلزم شناسایی پاسخ حافظه اپی ژنتیکی ناشی از تغذیه والدین است که مقدم بر توسعه فنوتیپ تغییر یافته در فرزندان است.
(2) اثرات تغذیه ای پدران بر روی علامت های اپيگنتیک پس از تولد
مطالعات در حال ظهور نشان داده اند که سطح تغذیه پدرانه می تواند بر رسوب چربی و علائم اپی ژنتیک فرزندان تأثیر بگذارد [31-34]. یک تحقیق اخیر با استفاده از موش نشان داده است که پیش دیابت پدرانه منجر به افزایش حساسیت به دیابت در فرزندان F1 با تغییرات مرتبط در بیان ژن لوزالمعده و متیلاسیون DNA مرتبط با سیگنالینگ انسولین می شود [35]. از همه مهمتر ، همپوشانی این تغییرات اپی ژنتیکی در جزایر پانکراس و اسپرم وجود دارد که نشان دهنده وراثت خط میکروب است. با این حال ، بیشتر این مطالعات ، اگرچه از نظر مفاهیم جذاب هستند ، اما در مقیاس ژنومی تحقیق محدود هستند و اغلب تغییرات اپی ژنتیکی ضعیف و تا حدی گذرا را در ارتباط با فنوتیپ های متابولیک خفیف در فرزندان نشان می دهند.
(iii) تغييرات اپيگنتيک نسل سوم بالقوه ايجاد رسوب چربي در پسران
انتقال پایدار اطلاعات اپی ژنتیک در نسل های چندگانه به خوبی در سیستم های گیاهی و C. elegans، اما اهمیت آن در پستانداران هنوز بسیار مورد بحث است [36، 37]. یک مبنای اپی ژنتیکی برای انتقال فنوتیپ ها از پدربزرگ و مادربزرگ در پاسخ به مواجهه با رژیم غذایی به خوبی ایجاد شده است، از جمله در گونه های دام [31]. تأثیرگذارترین مطالعاتی که اثرات انتقال اپی ژنتیکی را بر فنوتیپ فرزندان نشان میدهد، از نمونه موش آگوتی زرد زنده (Avy) استفاده کردهاند [38]. در این موش، قرار دادن یک رتروترانسپوزون در بالادست ژن آگوتی باعث بیان سازنده آن و در نتیجه رنگ زرد پوست و شروع چاقی بزرگسالان می شود. انتقال مادر از طریق خط زایا منجر به خاموش شدن بیان آگوتی به واسطه متیلاسیون DNA می شود که منجر به رنگ پوشش نوع وحشی و فنوتیپ لاغر فرزندان می شود [39، 40]. نکته مهم، مطالعات بعدی در این موش ها نشان داد که قرار گرفتن مادر در معرض اهداکنندگان متیل باعث تغییر رنگ پوشش می شود [41]. یک مطالعه انتقال یک فنوتیپ به نسل F3 و تغییرات در بیان تعداد زیادی ژن در پاسخ به محدودیت پروتئین در F0 را گزارش کرده است [42]. با این حال، تغییرات در بیان بسیار متغیر بود و ارتباط مستقیمی با تغییرات اپی ژنتیکی در این سیستم شناسایی نشد.
(iv) قرار گرفتن در معرض مستقیم افراد برای تغذیه بیش از حد در زندگی پس از زایمان
در حالی که بسیاری از مطالعات تغییرات اپی ژنتیکی مرتبط با رژیم را در مدل های حیوانی با استفاده از مناطق خاص منطقه نامشخص نشان داده اند، تجزیه و تحلیل چندین ژنوم انجام شده است. مطالعات اخیر بر روی تعیین اثر مستقیم اپی ژنتیکی رژیم های غذایی با چربی بالا / چاقی ناشی از رژیم غذایی در موش های بالغ با استفاده از بیان ژن ژنوم و تجزیه و تحلیل متیلاسیون DNA [43] متمرکز شده است. در این مطالعه، 232 به طور متفاوتی متوله شده (DMRs) در آديپوسیت ها از موش های کنترل و پرورش دهنده با چربی بالا شناخته شد. مهمتر از آن، مناطق مربوطه انسان برای DMR های موش نیز در بافت چربی از جمعیت چاق و لاغر متhylly متhylized شده، در نتیجه برجسته حفاظت تکاملی قابل توجه این مناطق. این نتیجه بر اهمیت احتمال DMR های شناسایی شده در تنظیم هومیوستاز انرژی در پستانداران تاکید دارد.
مطالعات انسانی
بر اساس شواهدی از مطالعات حیوانی و با در دسترس بودن ابزارهای مقرون به صرفه برای تجزیه و تحلیل ژنوم، گسترش سریع مطالعات اپی ژنوم در انسان صورت گرفته است. این مطالعات عمدتا بر شناسایی اختلافات خاص در محل در متيل شدن DNA متمرکز شده است که با فنوتيپهای متابوليک همراه است.
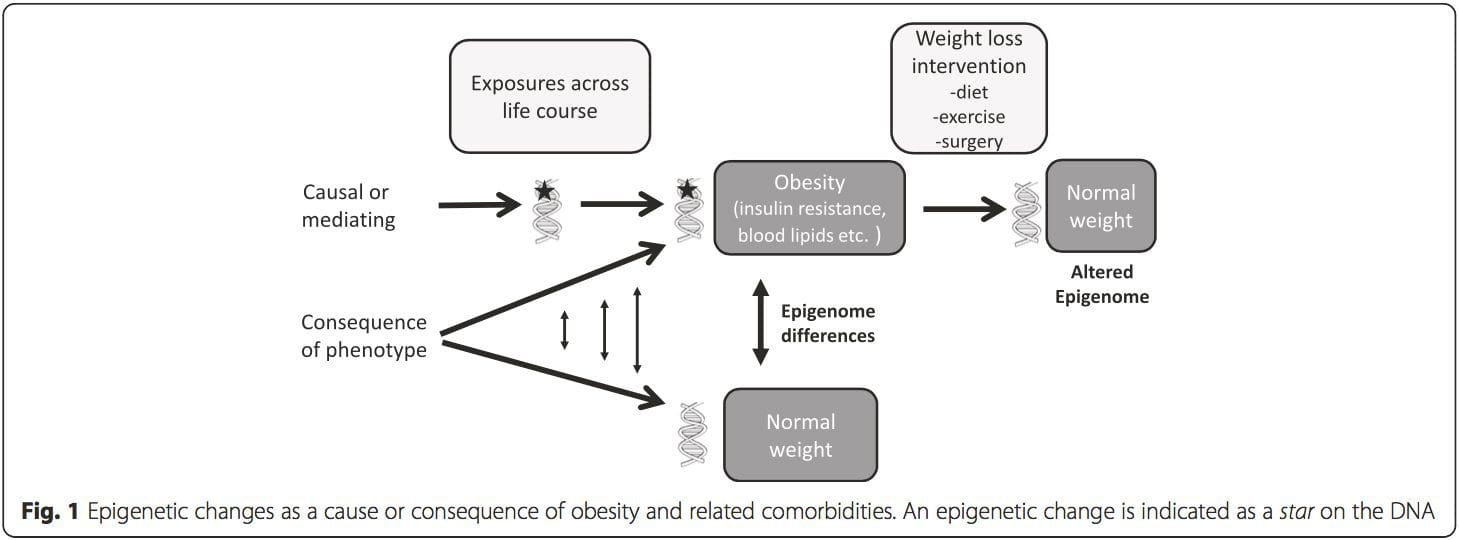
یک سوال کلیدی این است که تا چه حد تغییرات اپی ژنتیکی در ایجاد فنوتیپ متابولیک نقش داشته باشد، نه اینکه فقط یک دنباله آن باشد (شکل 1). برنامه ریزی اپی ژنتیک می تواند به رشد چاقی کمک کند، همچنین نقش مهمی در ریسک مشکلات قلبی عروقی و متابولیکی ایفا می کند. در مطالعات انسانی، اثبات علیت [44] دشوار است، اما می توان نتیجه گیری های متعددی را از تعدادی از شواهد انجام داد:
(i) مطالعات ارتباط ژنتیکی. پلیمورفیسمهای ژنتیکی که با افزایش خطر ابتلا به بیماریهای خاص مرتبط هستند، به طور پیشینی با ژنهای عامل مرتبط هستند. وجود متیلاسیون افتراقی در چنین مناطقی ارتباط عملکردی این تغییرات اپی ژنتیکی را در کنترل بیان ژن(های پروگزیمال) استنباط می کند. اثرات ژنتیکی قوی با اثر سیس وجود دارد که زیربنای بسیاری از تنوع اپی ژنتیکی است [7، 45]، و در مطالعات مبتنی بر جمعیت، روشهایی که از جانشینهای ژنتیکی برای استنباط نقش علّی یا واسطهای تفاوتهای اپی ژنوم استفاده میکنند [7، 46-48] استفاده شده است. . استفاده از اطلاعات ژنتیکی خانوادگی همچنین میتواند منجر به شناسایی مناطق کاندید بالقوه مسبب شود که متیلاسیون دیفرانسیل مرتبط با فنوتیپ را نشان میدهند [49].
(ii) زمان تغییرات اپی ژنتیکی. حضور علامت اپی ژنتیک قبل از توسعه یک فنوتیپ، یکی از ویژگی های اساسی مرتبط با علیت است. در مقابل، حضور علامت در ارتباط با چاقی، اما نه قبل از توسعه آن، می تواند برای حذف علیت استفاده شود، اما نقش احتمالی در آسیب شناسی مرتبط با چاقی را از بین نمی برد.
(iii) استدلال معتبر مکانیسم. این به تغییرات اپی ژنتیکی مربوط است که با بیان تغییرات ژن ها همراه با نقش تعیین شده در تنظیم فنوتیپ مورد علاقه مرتبط است. یکی از مثال های این است که ارتباط متیلاسیون در دو محل CpG در ژن CPT1A با سطوح تری گلیسیرید گردش خون [50]. CPT1A کدورت کارنیتین پالمیتویل ترانسفراز 1A، یک آنزیم با نقش مرکزی در متابولیسم اسید های چرب است، و این به شدت نشان می دهد که متفاوتی متیلاسیون این ژن ممکن است به علت تغییرات در غلظت تری گلیسرید پلاسما باشد.
مطالعات انجمن Epigenome-Wide: شناسایی بیومارکرهای اپی ژنتیک در سلامت متابولیک
تعدادی از تحقیقات اخیر بر روی بررسی ارتباط بین چاقی/بیماری های متابولیک و متیلاسیون DNA در سراسر ژنوم متمرکز شده اند (جدول 2). بزرگترین EWAS منتشر شده تاکنون، شامل 5465 نفر، 37 محل متیلاسیون در خون را شناسایی کرده است که با شاخص توده بدنی (BMI) مرتبط هستند، از جمله مکان هایی در CPT1A، ABCG1 و SREBF1 [51]. یک مطالعه در مقیاس بزرگ دیگر ارتباط ثابتی بین BMI و متیلاسیون در HIF3A در خون کامل و بافت چربی نشان داد [52]، یافته ای که تا حدی در مطالعات دیگر نیز تکرار شد [9، 51]. سایر ارتباطات اخیراً گزارش شده بین اقدامات مرتبط با چاقی و متیلاسیون DNA شامل (i) تفاوت متیلاسیون DNA بین لاغر و چاق افراد در LY86 در لکوسیت های خون [53] ؛ (ii) ارتباط بین متیلاسیون پروموتر PGC1A در خون کامل کودکان و چاقی 5 سال بعد [54]. (III) ارتباط بین نسبت دور کمر و باسن و متیلاسیون ADRB3 در خون [55]. و (IV) ارتباط بین BMI ، اقدامات توزیع چربی بدن و چندین سایت متیلاسیون DNA در بافت چربی [9 ، 56]. EWAS همچنین ارتباطاتی را بین سایت های متیلاسیون DNA و چربی های خون [55 ، 57-59] ، متابولیت های سرم [60] ، مقاومت به انسولین [9 ، 61] و T2DM [48 ، 62 ، 63] نشان داده است (جدول 2).
از این مطالعات، متیلاسیون تغییرات PGC1A، HIF3A، ABCG1 و CPT1A و RXRA [18] که قبلا توصیف شده است، به عنوان نشانگرهای زیستی مرتبط با سلامت متابولیک یا شاید پیش بینی کننده آن، متاثر از کاندیداهای قابل قبول برای نقش در توسعه بیماری متابولیک .
تعامل بین ژنوتیپ و Epigenome
تنوع اپی ژنتیکی به شدت تحتتاثیر تنوع ژنتیکی زیربنایی قرار میگیرد، با تخمین زده میشود که ژنوتیپ 20 تا 40 درصد از تغییرات را توضیح دهد [6، 8]. اخیراً، تعدادی از مطالعات شروع به ادغام دادههای متیلوم و ژنوتیپ برای شناسایی مکانهای صفت کمی متیلاسیون (meQTL) مرتبط با فنوتیپهای بیماری کردهاند. به عنوان مثال، در بافت چربی، یک meQTL همپوشانی با منبع خطر ژنتیکی BMI در یک عنصر تقویت کننده بالادست ADCY3 شناسایی شده است [8]. مطالعات دیگر همچنین همپوشانی هایی را بین چاقی شناخته شده و جایگاه های خطر T2DM و DMR های مرتبط با چاقی و T2DM شناسایی کرده اند [43، 48، 62]. متیلاسیون تعدادی از این DMRها نیز با تغذیه با چربی بالا در موش [43] و کاهش وزن در انسان [64] تعدیل شد. این نتایج یک پیوند جالب بین تغییرات ژنتیکی مرتبط با حساسیت به بیماری و ارتباط آنها با مناطقی از ژنوم را که در پاسخ به چالشهای تغذیهای دستخوش تغییرات اپی ژنتیکی میشوند، مشخص میکند که دلالت بر یک رابطه علّی دارد. ارتباط نزدیک بین تنوع ژنتیکی و اپی ژنتیکی ممکن است نشان دهنده نقش اساسی آنها در ایجاد تنوع فردی باشد [65، 66]. با این حال، در حالی که این یافته ها نشان می دهد که متیلاسیون DNA ممکن است واسطه اثرات ژنتیکی باشد، همچنین مهم است که در نظر بگیریم که هر دو فرآیند ژنتیکی و اپی ژنتیکی می توانند به طور مستقل بر روی ژن های یکسان عمل کنند. مطالعات دوقلو [8، 63، 67] میتوانند بینشهای مهمی ارائه دهند و نشان دهند که تفاوتهای بین فردی در سطوح متیلاسیون DNA عمدتاً از محیط غیرمشترک و تأثیرات تصادفی ناشی میشود، حداقل از اثرات محیطی مشترک، اما همچنین با تأثیر قابل توجه ژنتیکی. تغییر.
تأثیر محیط زیست قبل و بعد از زایمان بر روی Epigenome
محیط زیست قبل از زایمان: دو مطالعه که اخیراً منتشر شده است ، از جمعیت انسانی استفاده می کند که تغییرات طبیعی در تأمین مواد مغذی را تجربه کرده اند تا تأثیر تغذیه مادر قبل یا حین بارداری بر متیلاسیون DNA در فرزندان را بررسی کند. اولین مطالعه با استفاده از یک گروه مادر و کودک گامبیایی نشان داد که هر دو تغییر فصلی در مصرف متیل دهنده مادر در دوران بارداری و BMI مادر قبل از بارداری با تغییر متیلاسیون در نوزادان ارتباط دارد [68]. مطالعه دوم با استفاده از فرزندان بزرگسال از گروه گرسنگی گرسنگی هلندی برای بررسی اثر قرار گرفتن در معرض بارداری در یک دوره حاد سو und تغذیه شدید مادر بر متیلاسیون DNA ژن های درگیر در رشد و متابولیسم در بزرگسالی [69]. نتایج اهمیت زمان قرار گرفتن در معرض را در تأثیر آن بر اپی ژنوم برجسته کرد ، زیرا اثرات قابل توجه اپی ژنتیکی فقط در افرادی که در اوایل حاملگی در معرض قحطی قرار داشتند ، مشخص شد. نکته مهم ، تغییرات اپی ژنتیکی همراه با افزایش BMI است. با این حال ، نمی توان در این مطالعه مشخص کرد که آیا این تغییرات در اوایل زندگی وجود داشته اند یا نتیجه یک BMI بالاتر است.
سایر مطالعات اخیر شواهدی را ارائه داده اند که نشان می دهد تغذیه بیش از حد قبل از تولد و یک محیط مادر چاق یا دیابتی نیز با تغییرات متیلاسیون DNA در ژن های مربوط به رشد جنینی ، رشد و بیماری متابولیک در فرزندان ارتباط دارد [70-73]
در حالی که داده های انسانی کمیاب هستند، نشانه هایی وجود دارد که چاقی پدران می تواند منجر به تغییر متلاطم ژن های چاپ شده در نوزادان [74] شود، اثری است که به نظر می رسد که از طریق تغییرات اپی ژنتیکی در طی اسپرماتوژنز به دست می آید.
محیط زیست پس از زایمان: اپی ژنوم در طی رشد جنینی از نو تأسیس می شود و بنابراین ، به احتمال زیاد محیط قبل از تولد بیشترین تأثیر را روی اپی ژنوم دارد. با این حال ، اکنون مشخص شده است که تحت تأثیر طیف وسیعی از شرایط ، از جمله پیری ، قرار گرفتن در معرض سموم و تغییرات رژیم غذایی ، تغییراتی در اپی ژنوم بالغ ایجاد می شود. به عنوان مثال ، تغییرات متیلاسیون DNA در ژنهای متعدد در عضله اسکلتی و PGC1A در بافت چربی در پاسخ به رژیم غذایی پرچرب نشان داده شده است [75 ، 76]. مداخلات برای از دست دادن توده چربی بدن نیز با تغییر در متیلاسیون DNA همراه بوده است. مطالعات گزارش داده اند که پروفایل متیلاسیون DNA بافت چربی [43 ، 64] ، سلولهای تک هسته ای خون محیطی [77] و بافت عضلانی [78] در بیماران چاق قبلاً بیشتر به مشخصات افراد لاغر بدنبال کاهش وزن شباهت دارد. جراحی کاهش وزن همچنین تغییرات متیلاسیون مرتبط با بیماری کبد چرب غیر الکلی را تا حدی معکوس کرد [79] و در مطالعه دیگری منجر به هیپومتیلاسیون ژن های متعدد کاندید چاقی شد ، در مقایسه با چربی مطلق (احشایی) اثرات بارزتری در زیر جلدی داشت [64] . شواهد جمع آوری شده نشان می دهد که مداخلات ورزشی همچنین می تواند بر متیلاسیون DNA تأثیر بگذارد. بیشتر این مطالعات در افراد لاغر انجام شده است [80-82] ، اما یک مطالعه ورزشی در افراد چاق T2DM همچنین تغییراتی در متیلاسیون DNA را نشان داده است ، از جمله در ژن های درگیر در انتقال اسیدهای چرب و گلوکز [83]. تغییرات اپی ژنتیکی نیز با افزایش سن رخ می دهد ، و داده های اخیر نقش چاقی در افزایش آنها را نشان می دهد [9 ، 84 ، 85]. چاقی ، سن اپی ژنتیک بافت کبد را تسریع می کند ، اما بر خلاف یافته های توضیح داده شده در بالا ، این اثر پس از کاهش وزن قابل برگشت نیست [84].
در مجموع، شواهدی که در حمایت از ظرفیت تغذیه اپی ژن در بزرگسالان بیان شده است، نشان می دهد که ممکن است در زندگی پس از زایمان دخالت داشته باشد تا برنامه ریزی های متداول اپیزیونیک را مدون یا معکوس کند.
اندازه و اندازه اثر و تفاوت بین انواع بافت
تغییرات متیلاسیون DNA در ارتباط با چاقی یا ناشی از رژیم های غذایی یا مداخلات در سبک زندگی و کاهش وزن معمولاً ناچیز است (<15٪) ، اگرچه این بسته به فنوتیپ و بافت مورد مطالعه متفاوت است. به عنوان مثال ، تغییرات بیشتر از 20٪ در بافت چربی پس از کاهش وزن گزارش شده است [64] و ارتباط بین متیلاسیون HIF3A و BMI در بافت چربی بیشتر از خون بود [52].
نسبت بیولوژیکی تغییرات متیلاسیون نسبتا کوچک مورد سوال قرار گرفته است. با این حال، در بافت های متشکل از مخلوطی از انواع سلول، تغییر کوچک در متیلاسیون DNA ممکن است در واقع نشان دهنده تغییر قابل توجهی در یک کسر سلولی خاص است. ادغام داده های اپی ژنوم با داده های پروپکتروم و دیگر اپی ژنتیک، مانند تغییرات هیستون، مهم است، زیرا تغییرات متیلاسیون DNA کوچک ممکن است منعکس کننده تغییرات بزرگ در ساختار کروماتین باشد و می تواند با تغییرات وسیع تر در بیان ژن همراه باشد. زمینه ژنوم نیز باید مورد توجه قرار گیرد؛ تغییرات کوچک در یک عنصر نظارتی نظیر یک پروموتر، تقویتکننده، یا مقره ممکن است اهمیت عملکردی داشته باشد. در این راستا، DMR ها برای چاقی، و نیز مناطق تحت تاثیر قرار گرفتن در معرض گرسنگی قبل از زایمان و meQTL برای لوکوس صفت متابولیک مشاهده شده است که با هم ترکیب عناصر تقویت کننده [8، 43، 68]. شواهدی وجود دارد مبنی بر اینکه متیلاسیون DNA در مناطق وابسته به قحطی واقعا می تواند بر فعالیت فعال کننده [68] تأثیر بگذارد و از نقش تغییرات متیلاسیون ناشی از تغذیه در تنظیم ژن حمایت کند.
یک محدودیت عمده در بسیاری از مطالعات انسانی این است که علائم اپی ژنتیک اغلب در خون محیطی به جای بافت های مرتبط با متابولیسم ارزیابی می شود (شکل 2). ناهمگونی خون یک مسئله است، زیرا جمعیت های سلولی مختلف دارای علائم اپی ژنتیکی متمایز هستند، اما الگوریتم هایی برای تخمین ترکیب سلولی برای غلبه بر این مشکل توسعه یافته اند [86]. شاید مهمتر از آن، علائم اپی ژنتیک در سلول های خونی ممکن است لزوما وضعیت بافت های مورد علاقه اولیه را گزارش نکنند. با وجود این، مطالعات اخیر شواهد روشنی از رابطه بین علائم اپی ژنتیک در سلول های خونی و BMI ارائه کرده است. در مورد HIF3A که سطح متیلاسیون (مقدار بتا) در جمعیت مورد مطالعه از 0.14 تا 0.52 متغیر بود، افزایش 10 درصدی متیلاسیون با افزایش BMI به میزان 7.8 درصد همراه بود[52]. به همین ترتیب، یک تفاوت 10 درصدی در متیلاسیون PGC1A ممکن است تا 12 درصد تفاوت در توده چربی را پیش بینی کند [54].
نتیجه گیری
مطالعه نقش اپی ژنتیک در چاقی و بیماری متابولیک در سال های اخیر به سرعت در حال افزایش است و شواهد نشان می دهد که یک پیوند بین تغییرات اپيگنتیک و نتایج سلامت متابولیک در انسان وجود دارد. بیومارکرهای اپی ژنتیکی بالقوه مرتبط با چاقی و سلامت متابولیکی نیز از مطالعات اخیر پدید آمده است. اعتبارسنجی علامت های اپی ژنتیک در گروه های مختلف، این واقعیت که چندین علامت در ژن ها با عملکرد قابل اعتماد در چاقی و توسعه T2DM و همچنین همپوشانی علامت های اپی ژنتیک با چاقی شناخته شده و لکنت های ژنتیکی T2DM یافت می شود، شواهدی را ثابت می کند که این انجمن ها واقعی تا به امروز کشمکش بسیار دشوار است. با این وجود، صرف نظر از این که انجمن ها علت هستند، علامت های اپی ژنتیکی شناسایی شده ممکن است همچنان به عنوان نشانگرهای زیست شناختی مربوط باشد چاقی و خطر ابتلا به بیماری متابولیک.
اندازه اثرات در بافتهای به راحتی در دسترس مانند خون کوچک است اما به نظر می رسد با وجود تغییر در قومیت ، نوع بافت و روش های تجزیه و تحلیل ، قابل تجدید باشد [51]. همچنین ، حتی تغییرات کوچک متیلاسیون DNA نیز ممکن است دارای اهمیت بیولوژیکی باشند. یک رویکرد "اومیک" یکپارچه در گشودن بیشتر فعل و انفعالات پیچیده بین اپی ژنوم ، رونویسی ، ژنوم و سلامت متابولیکی بسیار مهم است. مطالعات طولی ، که به طور ایده آل چندین نسل را در بر می گیرد ، برای ایجاد روابط علی ضروری است. در آینده می توان انتظار چنین مطالعات بیشتری را داشت ، اما این کار زمان بر است.
در حالیکه مطالعات حیوانی همچنان اثری از زندگی زودهنگام را نشان می دهد تغذیه قرار گرفتن در معرض اپی ژنوم و سلامت متابولیک فرزندان، داده های انسانی هنوز محدود است. با این حال، مطالعات اخیر شواهد روشنی را ارائه کرده اند که قرار گرفتن در معرض تغذیه نامناسب در طول دوره های خاص رشد پیش از تولد با تغییرات متیلاسیون در فرزندان مرتبط است و بنابراین پتانسیل تأثیرگذاری بر فنوتیپ بالغ را دارد. مطالعات حیوانی برای تأیید یافتههای انسانی در یک محیط کنترلشدهتر، کمک به تعیین اینکه آیا تغییرات متیلاسیون شناساییشده تأثیری بر سلامت متابولیک دارد یا خیر، و کشف مکانیسمهای زیربنایی این مقررات اپی ژنتیکی بین نسلی/فرنسلی مهم خواهد بود. شناسایی مکانیسمهای علّی زیربنای پاسخهای حافظه متابولیک، نحوه انتقال اثرات فنوتیپی به نسلهای متوالی، میزان تأثیر و پایداری صفت منتقلشده، و شناسایی یک زمینه تکاملی فراگیر و متحدکننده نیز سؤالات مهمی هستند که باید به آنها پرداخته شود. . دومی اغلب با فرضیه پاسخ تطبیقی پیشبینیکننده، یعنی پاسخی به یک محیط پیشبینیشده در آینده که تناسب اندام جمعیت را افزایش میدهد، محصور میشود. با این حال، این فرضیه به طور فزاینده ای مورد تردید قرار گرفته است زیرا شواهد محدودی برای افزایش تناسب اندام در مراحل بعدی زندگی وجود دارد [87].
به طور خلاصه، نتایج امیدوار کننده است، به عنوان تغییرات اپی ژنتیک با سلامت متابولیسم بزرگسالان مرتبط است و آنها به عنوان یک واسطه بین تغذیه تغذیه قبل از زایمان و افزایش خطر ابتلا به نتایج بدنی متابولیسم ناشی از آن عمل می کنند. علائم اپی ژنتیک جدید شناسایی شده اند که با اندازه گیری های سلامت متابولیکی همراه هستند. ادغام لایه های مختلف اطلاعات ژنتیکی، حمایت بیشتر از روابط علی را افزوده است، و مطالعات بیشتری نشان داده اند که اثرات محیط زیست قبل و بعد از زایمان بر روی پروتئین و سلامت وجود دارد. در حالی که بسیاری از سوالات مهم همچنان باقی می مانند، پیشرفت های روش شناختی اخیر، انواع مطالعات مبتنی بر جمعیت مبتنی بر بزرگ را که برای حل شکاف دانش لازم است، فراهم می کند. دهه آینده وعده داده است که یک دوره فعالیت اصلی در این منطقه تحقیقاتی مهم باشد.
Susan J. van Dijk1، Ross L. Tellam2، Janna L. Morrison3، Beverly S. Muhlhausler4,5،1 و Peter L. MolloyXNUMX *
رقابت منافع
نویسندگان اعلام می کنند که آنها منافع رقابتی ندارند.
مشارکت نویسندگان
همه نویسندگان به تهیه پیش نویس و بازنگری انتقادی این دست نویس کمک کردند و همه نویسندگان مقاله نهایی را خواندند و تایید کردند.
اطلاعات نویسندگان
بورلی S. Muhlhausler و پیتر L. مولوی اخیر نویسندگان مشترک هستند.
سپاسگزاریها
این کار توسط کمک مالی از صندوق تأسیسات علمی و صنعت (Grant RP03-064) پشتیبانی شده است. JLM و BSM توسط شورای ملی برنامه های توسعه شغلی شورای ملی بهداشت و درمان (JLM، APP1066916، BSM، APP1004211) پشتیبانی می شوند. ما از Lance Macaulay و Sue Mitchell برای خواندن انتقادی و نظرات در مورد دستنوشته تشکر می کنیم.
جزئیات نویسنده
1CSIRO Food and Nutrition Flagship، صندوق پستی 52، نورث راید، NSW 1670، استرالیا. 2CSIRO Agriculture Flagship، 306 Carmody Road، St Lucia، QLD 4067، استرالیا. 3خاستگاه اولیه گروه تحقیقاتی سلامت بزرگسالان، دانشکده داروسازی و علوم پزشکی، موسسه تحقیقات بهداشتی سانسوم، دانشگاه استرالیای جنوبی، صندوق پستی 2471، آدلاید، SA 5001، مرکز تحقیقاتی استرالیا4FOODplus، پردیس وایت، دانشگاه آدلاید، PMB 1, Glen Osmond, SA 5064, استرالیا. 5 مؤسسه تحقیقاتی سلامت زنان و کودکان، خیابان کینگ ویلیام 72، آدلاید شمالی، SA 5006، استرالیا.
جای خالی
منابع:
1. چه کسی WHO | اضافه وزن و چاقی www.who.int/gho/ncd/
risk_factors / اضافه وزن / en / index.html. دسترسی به 29 ژانویه 2015.
2 Visscher PM، براون MA، McCarthy MI، یانگ J. پنج سال از کشف GWAS.
من جی هوم ژنت هستم. 2012؛ 90:7:24.
3 لک AE، Kahali B، Berndt SI، عدالت AE، Pers TH، روز FR، و غیره. ژنتیک
مطالعات شاخص توده بدنی بینش جدیدی را برای زیست شناسی چاقی ایجاد می کند. طبیعت
2015 ؛ 518: 197 206.
4. لینگ سی، دل گورا اس، لوپی آر، رن تی، گرانهال سی، لوتمن اچ، و همکاران.
تنظیم اپيگنتيک PPARGC1A در جزایر ديابتی 2 انسان و
اثر بر ترشح انسولین دیابت شناسی 2008؛ 51:615~22.
5 وان دیک SJ، مولوی PL، وینلی هور، موریسون JL، Muhlhausler BS. اپی ژنتیک
و چاقی انسان Int J Obes (لند). 2015؛ 39:85 97.
6 AL، Pan H، چن L، Ong ML، Dogra S، Wong J، و همکاران. اثر از
ژنوتيپ و در محيط رحم در تغييرات درون فردي در نوزادان
متیلوم های DNA ژنوم Res. 2014؛ 24:1064 74.
7 Olsson AH، Volkov P، Bacos K، Dayeh T، Hall E، Nilsson EA، et al. Genomewide
ارتباط بین نفوذ تنوع ژنتیکی و اپی ژنتیکی
بیان mRNA و ترشح انسولین در جزایر پانکراس انسانی. PLoS
جنت 2014؛ 10: e1004735.
8 Grundberg E، Meduri E، Sandling JK، Hedman AK، Keildson S، Buil A، و همکاران.
تجزیه و تحلیل جهانی تغییرات متیلاسیون DNA در بافت چربی از دوقلوها
لینک های مرتبط با انواع مرتبط با بیماری در عناصر قانونی دفاعی را نشان می دهد.
من جی هوم ژنت هستم. 2013؛ 93:876:90.
9 Ronn T، Volkov P، Gillberg L، Kokosar M، Perfilyev A، Jacobsen AL، و همکاران.
تاثیر سن، BMI و سطح HbA1c بر روی ژنوم DNA گسترده
متیلاسیون و الگوهای بیان mRNA در بافت چربی انسان
و شناسایی بیومارکرهای اپی ژنتیک در خون. Hum Mol Genet.
2015 ؛ 24: 3792 813.
10 Waterland RA، میچلز KB. اپیدمیولوژی اپیزیونیک رشد
فرضیه ریشه ها Annu Rev Nutr. 2007؛ 27:363 88.
11 McMillen IC، Rattanatray L، Duffield JA، موریسون JL، MacLaughlin SM، Gentili
S و همکاران ریشه های اولیه چاقی بعد: مسیرها و مکانیسم ها. مشاوره
Exp Med Biol. 2009؛ 646:71~81.
12 Ravelli A، Van der Meulen J، Michels R، Osmond C، Barker D، Hales C، et al.
تحمل گلوکز در بزرگسالان پس از مواجهه با قحطی قبل از زایمان. لانست
1998 ؛ 351: 173 7.
13 McMillen IC، MacLaughlin SM، Muhlhausler BS، Gentili S، Duffield JL،
موریسون جی. ال. ریشه های رشد سلامت و بیماری بالغ: نقش
تغذیه سالم و جنین. کلین عمومی کالج داروسازی Toxicol.
2008 ؛ 102: 82 9.
14 ژانگ S، Rattanatray L، McMillen IC، Suter CM، موریسون JL. Periconceptional
تغذیه و برنامه ریزی اولیه زندگی چاقی و یا بیماری. پروگ
Biophys Mol Biol. 2011؛ 106:307 14.
15 بوت S، Levin BE، Ozanne SE. کنترل های تعامل ژن و محیط زیست
انرژی و هومیوستاز گلوکز و ریشه های رشد چاقی.
Physiol Rev. 2015؛ 95:47~82.
16 Borengasser SJ، Zhong Y، Kang P، Lindsey F، Ronis MJ، Badger TM و همکاران.
چاقی مادر باعث افزایش تمایز بافت های سفید چربی می شود و تغییر می کند
DNA متیلاسیون در ژنوم در فرزندان موش صحرایی. غدد درون ریز
2013 ؛ 154: 4113 25.
17 Gluckman PD، Lillycrop KA، Vickers MH، Pleasants AB، Phillips ES، Beedle AS،
و همکاران پلاستیک متابولیک در طول توسعه پستانداران به طور مستقیم است
وابسته به وضعیت تغذیه زودرس. Proc Natl Acad Sci USA A.
2007 ؛ 104: 12796 800.
18 Godfrey KM، Sheppard A، Gluckman PD، Lillycrop KA، Burdge GC، McLean C،
و همکاران متیلاسیون پروموتر ژن اپیژنیک در هنگام تولد همراه است
چاقی بعدی کودک دیابت. 2011؛ 60: 1528~34.
19 McMillen IC، Adam CL، Muhlhausler BS. ریشه های اولیه چاقی:
برنامه ریزی سیستم تنظیم اشتها جی فیزیول. 2005؛ 565 (Pt 1): 9.
20 Begum G، Stevens A، Smith EB، Connor K، Challis JR، Bloomfield F و همکاران.
تغییرات اپی ژنتیکی در مسیرهای تنظیم کننده انرژی هیپوتالاموس جنین
همراه با ناتوانی در مادران و دوزندگی. FASEB J.
2012 ؛ 26: 1694 703.
21 جی جی جی، لیانگ QX، هو ی، هان ZM، شاتن هان، سان QY و همکاران. چاقی مادر
و دیابت ممکن است دگرگونی متیلاسیون DNA در اسپرم ها را ایجاد کند
فرزندان در موش. تجدید Biol Endocrinol. 2014؛ 12: 29.
22 جوس C، پری لی، لامبرت Langlais س، مورین AC، Averous J، برات A، و غیره.
ناقل تغذیه پری ناتال بر متیلاسیون و بیان لپتین تاثیر می گذارد
ژن در بزرگسالان: معیار برای درک سندرم متابولیک.
FASEB J. 2011؛ 25:3271~8.
23 Lan X، Cretney EC، Kropp J، Khateeb K، Berg MA، Penagaricano F، و همکاران.
رژیم غذایی مادر در دوران بارداری موجب بیان ژن و DNA می شود
تغییرات متیلاسیون در بافت جنین در گوسفند. جت جلو 2013؛ 4: 49.
24 Li CC، Young PE، Maloney CA، Eaton SA، Cowley MJ، Buckland ME، et al.
چاقی مادر و دیابت موجب نقص های متابولیسم پنهان و
تغییرات اپی ژنتیکی گسترده در موش های ایزوژنیک اپی ژنتیک 2013؛ 8:602.
25 Lillycrop KA، Phillips ES، جکسون AA، Hanson MA، Burdge GC. پروتئین غذایی
محدودیت موش های حامله و الکل و مکمل های اسید فولیک مانع می شود
اصلاح اپی ژنتیک بیان ژن کبدی در پسران. J Nutr
2005 ؛ 135: 1382 6.
26 Radford EJ، Ito M، Shi H، Corish JA، Yamazawa K، Isganaitis E، و همکاران. در دوران جنینی
اثرات در فقر آهن رحم متابولیسم بالغ اسپرم متورم است
و متابولیسم بین نسلی. علوم پایه. 2014؛ 345 (80): 1255903.
27 Suter M، Bocock P، Showalter L، Hu M، Shope C، McKnight R، و همکاران.
Epigenomics: قرار گرفتن در معرض رژیم غذایی رژیم غذایی با رژیم غذایی بالا در رژیم های غذایی رحم
بیان ژن محیطی محیطی در افراد غیر انسانی. FASEB J.
2011 ؛ 25: 714 26.
28 سوتر MA، م ج، Vuguin PM، Hartil K، Fiallo A، Harris RA، و غیره. در دوران جنینی
قرار گرفتن در معرض یک رژیم غذایی با چربی مادری، کد هیستون اپی ژنتیکی را تغییر می دهد
مدل موش Am J Obs Gynecol. 2014؛ 210:463 e1 e463.
29 حرف مفت DN، فو Q، کالاوی CW، مک نایت RA، McMillen IC، راس MG، و همکاران.
اپی ژنتیک چاقی برنامه ریزی شده: تغییر در IGR IGRX کبدی IGF1
بيان ژن mRNA و ساختار هيستون در سريع و بعد از تولد تاخير دارد
رشد رو به افزایش است Am J Physiol Gastrointest Liver Physiol.
2010؛ 299: G1023 9.
30 Sandovici I، اسمیت NH، Nitert MD، Ackers-جانسون M، Uribe-لوئیس S، Ito Y،
و همکاران رژیم مادری و پیری کنترل اپی ژنتیک پرومتئورنبرگ را تغییر می دهد
تعامل در ژن Hnf4a در جزایر پانکراس موش صحرایی. Proc Natl
Acad Sci US A. 2011؛ 108:5449~54.
31 Braunschweig M، Jagannathan V، Gutzwiller A، Bee G. تحقیقات در مورد
پاسخ اپی ژنتیک ترانس ژنراتور را به صورت نر در خوک های F2 پایین می آورد. PLoS
یکی 2012؛ 7، e30583.
32 Carone BR، Fauquier L، Habib N، Shea JM، Hart CE، Li R، et al. پدرانه
برنامه ریزی متابولیکی محیط زیست ترانس ژنراتور ایجاد شده است
بیان ژن در پستانداران سلول. 2010؛ 143:1084 96.
33 Ost A، Lempradl A، Casas E، Weigert M، Tiko T، Deniz M، و همکاران. رژیم پدرانه
وضعیت کروماتین فرزندان و چاقی بین نسلی را تعریف می کند. سلول.
2014 ؛ 159: 1352 64.
34. مارتنز دی، پنتینات تی، ریب اس، داویود سی، بلوکس وی دبلیو، سبری جی، و همکاران. در دوران جنینی
سوء تغذیه در موش های نر برنامه متابولیسم لیپید های کبدی را در مرحله دوم تولید می کند
فرزندان شامل متلاشی شدن DNA Lxra. سلول متاب.
2014 ؛ 19: 941 51.
35 Wei Y، Yang CR، Wei YP، Zhao ZA، Hou Y، Schatten H، et al. پدرانه
ناشی از ارثی ترانس ژنراتوری حساسیت به دیابت در
پستانداران Proc Natl Acad Sci US A. 2014؛ 111:1873~8.
36 Grossniklaus U، کلی WG، کلی B، فرگوسن اسمیت AC، Pembrey M، Lindquist
S. توزیع اپی ژنتیک ترانس نسبی: چقدر مهم است؟ Nat Rev
ژنت 2013؛ 14:228:35.
37 Pembrey M، Saffery R، Bygren LO. پاسخ های انسانی انسان به
تجربه زود هنگام زندگی: تاثیر بالقوه بر توسعه، سلامت و
تحقیقات زیست پزشکی جی مد ژنت. 2014؛ 51:563 72.
38 Wolff GL، Kodell RL، Moore SR، Cooney CA. اپی ژنتیک مادر و متیل
مکمل ها بر بیان ژن آگوتای در Avy / موش تاثیر می گذارد. FASEB J.
1998 ؛ 12: 949 57.
39 Jirtle RL، Skinner MK. Epigenomics محیطی و حساسیت به بیماری.
Nat Rev Genet. 2007؛ 8:253 62.
40 مورگان HD، Sutherland HG، مارتین DI، Whitelaw E. ارگانیک اپیزیونیک در
مکان آگوتی در موش. نات ژنت. 1999؛ 23:314~8.
41 کرولی JE، Suter CM، Beckman KB، Martin DI. اپی ژنتیک گیاهی
اصلاح موش آلوئه مغز توسط مکمل های تغذیه ای. Proc
Natl Acad Sci US A. 2006؛ 103:17308-12.
42 Hoile SP، Lillycrop KA، Thomas NA، Hanson MA، Burdge GC. پروتئین غذایی
محدودیت در حاملگی F0 در موش باعث تغییرات ترانس ژنراتور در
ترانسکتوموم کبدی در پسران زن. PLoS یکی. 2011؛ 6، e21668.
43 Multhaup ML، Seldin MM، Jaffe AE، Lei X، Kirchner H، Mondal P، و همکاران. Mousehuman
تجزیه و تحلیل اپی ژنتیکی تجربی باعث نگرانی از اهداف رژیم غذایی و
مسئولیت ژنتیکی برای فنوتیپ های دیابتی سلول متاب. 2015؛ 21:138.
44 میشلز KB، Binder AM، Dedeurwaerder S، Epstein CB، Greally JM، Gut I، و همکاران.
توصیه هایی برای طراحی و تجزیه و تحلیل گسترده اپی ژنوم
مطالعات انجمن روشهای Nat 2013؛ 10:949 55.
45. Dayeh TA, Olsson AH, Volkov P, Almgren P, Rânn T, Ling C. Identification of
CpG-SNPs مرتبط با دیابت نوع 2 و متیلاسیون DNA دیفرانسیل
در جزایر پانکراس انسان دیابت شناسی 2013؛ 56:1036~46.
46 Relton CL، Davey Smith G. تصادفي ماندلايی Epigenetic دو مرحلهای: a
استراتژی برای ایجاد نقش علمی فرایندهای اپی ژنتیکی در مسیرها
به بیماری Int J Epidemiol. 2012؛ 41:161.
47 لیو ی، آرایی مژده، Padyukov L، Fallin MD، Hesselberg E، Runarsson A، و غیره.
داده های ارتباطی گسترده Epigenome نشان می دهد که متیلاسیون DNA به عنوان یک
واسطه خطر ژنتیک در آرتریت روماتوئید. Nat Biotechnol.
2013 ؛ 31: 142 7.
48 یوان W، Xia Y، Bell CG، اما I، Ferreira T، Ward KJ و همکاران. یکپارچه
آنالیز اپی ژنومیک برای لوکوس های حساس به دیابت نوع 2 در مونوزیگوت
دوقلوها. Nat Commun. 2014؛ 5: 5719.
49 Nitert MD، Dayeh T، Volkov P، Elzzyri T، Hall E، Nilsson E، و همکاران. تاثیر یک
مداخله ورزشی بر روی متیلاسیون DNA در عضله اسکلتی از درجه اول
بستگان بیماران مبتلا به دیابت نوع 2. دیابت. 2012؛ 61:3322~32.
50. Gagnon F، A'ssi D، Carri A، Morange PE، Trâgou't DA. اعتبار سنجی قوی از
ارتباط سطح متیلاسیون در لوپوس CPT1A با سطح پلاسمای لیپید.
J Lipid Res. 2014؛ 55: 1189 91.
51 Demerath EW، Guan W، Grove ML، Aslibekyan S، Mendelson M، Zhou YH،
و همکاران BMI، تغییر BMI، و
دور کمر در بزرگسالان آفریقایی آمریکایی چندین تکرار را مشخص می کند
جایگاه. هوم مول ژنت. 2015:ddv161.
52. Dick KJ, Nelson CP, Tsaprouni L, Sandling JK, A’ssi D, Wahl S, et al. DNA
متیلاسیون و شاخص توده بدن: تجزیه و تحلیل گسترده ژنوم. لانست
2014 ؛ 6736: 1 9.
53 سو S، Zhu H، Xu X، Wang X، Dong Y، Kapuku G، و همکاران. متیلاسیون DNA از
ژن LY86 با چاقی، مقاومت به انسولین و
التهاب دوقلو رس هوم ژنه. 2014؛ 17:183 91.
54 Clarke-Harris R، Wilkin TJ، Hosking J، Pinkney J، Jeffery AN، Metcalf BS، و همکاران.
PGC1؟ متیلاسیون پروموتر در خون در 5 تا 7 سالگی، چاقی را پیش بینی می کند
9 تا 14 سال (EarlyBird 50). دیابت. 2014؛ 63:2528~37.
55 Guay SP، Brisson D، Lamarche B، Biron S، Lescelleur O، Biertho L، و همکاران.
DNA متیلاسیون پروتئین ADRB3 در خون و چربی احشایی
بافت با اختلالات متابولیکی در مردان ارتباط دارد. Epigenomics
2014 ؛ 6: 33 43.
56 Agha G، Houseman EA، Kelsey KT، Eaton CB، Buka SL، Loucks EB. چاقی است
همراه با مشخصات متیلاسیون DNA در بافت چربی. Int J اپیدمیول.
2014: 1 11.
57 ایروین م. ر.، زهی د، جوئن ر.، مندلسون م، اسلیبکن S، کلاوس س. و همکاران.
بررسی ارتباط کلی Epigenome در مورد چربی خون ناشتا در ژنتیک
داروهای کاهنده چربی و مطالعه شبکه رژیم غذایی جریان. 2014؛ 130:565~72.
58 Frazier-Wood AC، Aslibekyan S، Absher DM، Hopkins PN، Sha J، Tsai MY، و همکاران.
متیلاسیون در لوپوس CPT1A همراه با زیر فرایند لیپوپروتئین همراه است
پروفایل ها J Lipid Res. 2014؛ 55:1324 30.
59 Pfeifferm L، Wahl S، Pilling LC، Reischl E، Sandling JK، Kunze S و همکاران. DNA
متيل شدن ژن هاي مرتبط با چربي باعث کاهش سطح چربي خون مي شود. Circ Cardiovasc
جنت 2015
60. پترسن AK، زایلینگر اس، کاستنمولر جی، رومیش-مارگل دبلیو، بروگر ام، پیترز
A و همکاران Epigenetics با متابولومیکس همراه است
مطالعه با صفات متابولیک سرم خون هوم مول ژنت. 2014؛ 23:534 45.
61 Hidalgo B، Irvin MR، Sha J، Zhi D، Aslibekyan S، Absher D، et al. Epigenomewide
بررسی میزان مصرف روزانه گلوکز، انسولین و HOMA-IR
در ژنتیک داروهای کاهش چربی و رژیم غذایی رژیم غذایی. دیابت.
2014 ؛ 63: 801 7.
62. دایه تی، ولکوف پی، سالاس، هال ای، نیلسون ای، اولسون AH، و همکاران. در سطح ژنومی
تجزیه و تحلیل متیلاسیون DNA از جزایر پانکراس انسان از نوع دیابتی نوع 2
و اهداکنندگان غیر دیابتی ژنهای نامزدی که انسولین را تحت تاثیر قرار می دهند، شناسایی می کنند
ترشح PLoS جنت 2014؛ 10، e1004160.
63 Nilsson E، Jansson PA، Perfilyev A، Volkov P، Pedersen M، Svensson MK، و همکاران.
متیلاسیون DNA تغییر یافته و بیان دیفرانسیل ژن هایی که تحت تاثیر قرار می گیرند
متابولیسم و التهاب در بافت چربی از افراد با نوع 2
دیابت. دیابت. 2014؛ 63: 2962 76.
64 بنتون MC، Johnstone A، Eccles D، هارمون B، Hayes MT، Lea RA، و همکاران. تجزیه و تحلیل متیلاسیون DNA در بافت چربی انسان نشان می دهد تغییر دیفرانسیل از ژن های چاقی قبل و بعد از بای پس معده و وزن
ضرر - زیان. ژن. 2015؛ 16:1:21.
65 Bateson P، Gluckman P. پلاستیک و استحکام در توسعه و
سیر تکاملی. Int J Epidemiol. 2012؛ 41:219 23.
66 Feinberg AP، Irizarry RA، Feinberg AP، Irizarry RA. تکامل در سلامت و
طب مکمل و جایگزین Sackler: تنوع اپی ژنتیک تصادفی به عنوان یک رانندگی
نیروی توسعه، انطباق تکاملی و بیماری است. Proc Natl Acad
Sci US A. 2010; 107 (Suppl): 1757~64.
67 مارتینو دی، لک یج، گوردون ل، الیکینن م، ماریو کریکشکان، سافرری و همکاران.
تجزیه و تحلیل طولی، ژنوم در سطح متیلاسیون DNA در دوقلوها از زمان تولد
به ماه های 18، تغییرات سریع اپی ژنتیکی در زندگی زودرس و مشخصه های متفاوتی را نشان می دهد
اثرات عدم انطباق Genome Biol. 2013؛ 14: R42.
68 Tobi EW، Goeman JJ، Monajemi R، Gu H، Putter H، Zhang Y، et al. DNA
امضا های متیلاسیون در ارتباط با رشد قحطی روزانه است
متابولیسم Nat Commun. 2014؛ 5: 5592.
69 Dominguez-Salas P، Moore SE، Baker MS، Bergen AW، Cox SE، Dyer RA و همکاران.
تغذیه مادر در مفهوم ملاک سازی متیلاسیون DNA انسان است
معمای متاستاز Nat Commun. 2014؛ 5: 3746.
70 Quilter CR، Cooper WN، Cliffe KM، Skinner BM، Prentice PM، Nelson L، et al.
تأثیر الگوهای متیلاسیون فرزندان مادران دیابتی حاملگی
محدودیت رشد و رشد داخل رحمی نشان می دهد ژن های مشترک و
مسیرهای مرتبط با خطر دیابت نوع 2 بعدی FASEB J. 2014: 1�12.
71 Morales E، Groom A، Lawlor DA، Relton CL. امضاهای متیلاسیون DNA در
خون بند ناف همراه با افزایش وزن مادر بارداری: نتایج از
همکار ALSPAC. BMC Res Notes 2014؛ 7: 278.
72 Ruchat SM، Houde AA، Voisin G، St-Pierre J، Perron P، Baillargeon JP، et al.
دیابت حاملگی اپیزیونیک عمدتا بر ژنها تاثیر می گذارد
در بیماری های متابولیک نقش دارد. اپی ژنتیک 2013؛ 8:935 43.
73 لیو X، چن Q، Tsai HJ، وانگ G، هنگ X، Zhou Y، و غیره. مادری
شاخص توده بدنی پیش برده و DNA خون بند ناف فرزندان
متیلاسیون: کشف علل زودهنگام بیماری. Environ Mol
جهش زا. 2014؛ 55:223 30.
74 Soubry A، Murphy SK، وانگ F، هوانگ Z، Vidal AC، Fuemmeler BF، و همکاران.
نوزادان چاق والدین الگوهای متیلاسیون DNA را در
ژن های حک شده Int J Obes (لند). 2015؛ 39:650±7.
75. جاکوبسن SC، برنز سی، بورک-جنسن جی، ریبل-مدسن آر، یانگ بی، لارا ای، و همکاران.
اثرات کوتاه مدت چاقی بیش از حد چربی بر DNA ژنوم
متیلاسیون در ماهیچه اسکلتی مردان جوان سالم. دیابت
2012 ؛ 55: 3341 9.
76. Gillberg L، Jacobsen SC، Rânn T، Brâns C، Vaag A. PPARGC1A DNA
متیلاسیون در بافت چربی زیر جلدی در افراد کم وزن هنگام تولد
تاثیر 5 روز تغذیه بیش از حد با چربی بالا متابولیسم. 2014؛ 63:263 71.
77 هوانگ YT، Maccani JZJ، هاولی NL، Wing RR، Kelsey KT، McCaffery JM.
الگوهای اپيگنتيک در نگهدارندگان کاهش وزن موفق: يک مطالعه آزمايشي. اینتر ج
Obes (لند). 2015؛ 39:865~8.
78. Barres R، Kirchner H، Rasmussen M، Yan J، Kantor FR، Krook A، Nâslund E،
Zierath JR. کاهش وزن پس از عمل جراحی بای پس معده در چاقی انسان
متیلاسیون پروموتر را بازسازی می کند. Cell Rep. 2013: 1~8.
79. Ahrens M، Ammerpohl O، von Schónfels W، Kolarova J، Bens S، Itzel T، و همکاران.
تجزیه و تحلیل متیلاسیون DNA در بیماری کبد چرب غیر کلاسیک نشان می دهد
امراض اختصاصی بیماری و تجدید ساختار پس از جراحی بارداری.
سلول متاب. 2013؛ 18:296 302.
80 Voisin S، Eynon N، Yan X، اسقف DJ. آموزش تمرین و متیلاسیون DNA
در انسان Acta Physiol (Oxf). 2014؛ 213:39 59.
81. لیندهولم می، مارابیتا اف، گومز-کابررو دی، راندکویست اچ، اکستروم تی جی،
تگنر جی و همکاران تجزیه و تحلیل یکپارچه برنامه ریزی مجدد هماهنگ را نشان می دهد
از epigenome و ترانسکتوموم در عضله اسکلت انسان پس از
آموزش. اپی ژنتیک 2014؛ 9:1557 69.
82. Denham J، O'Brien BJ، Marques FZ، Charchar FJ. تغییرات در لکوسیت
متيلوم و اثر آن بر ژن هاي مرتبط با قلب و عروق پس از ورزش.
J Applis Physiol. 2014: jap.00878.2014.
83 Rowlands DS، صفحه RA، Sukla WR، Giri M، Ghimbovschi SD، Hayat I، و همکاران.
شبکه های مجتمع چند omic DNA متیلاسیون و miRNA را با یکدیگر متصل می کنند
پلاستیک عضله اسکلتی به ورزش مزمن در چاقی دیابت نوع 2.
فیزیول ژنومیکس. 2014؛ 46:747 65.
84 Horvath S، Erhart W، Brosch M، Ammerpohl O، فون Schonfels W، Ahrens M،
و همکاران چاقی، پیری اپی ژنتیک کبد انسان را تسریع می کند. Proc Natl Acad
علمی 2014؛ 111: 15538~43.
85. Almón MS، Nilsson EK، Jacobsson JA، Kalnina I، Klovins J، Fredriksson R، و همکاران.
تجزیه و تحلیل ژنوم نشان می دهد نشانگر متیلاسیون DNA که با متفاوت است
هم سن و هم چاقی ژن. 2014.؛ 548:61
86 Houseman EA، Molitor J، Marsit CJ. اصلاح مخلوط سلول های مرجع
در تجزیه و تحلیل داده های متیلاسیون DNA. بیوانفورماتیک. 2014؛ 30:1431.
87 Wells JC ارزیابی انتقادی از فرضیه پیشنهادی انطباق پذیری.
Int J Epidemiol. 2012؛ 41:229:35.
88 ویلیامز-ویس O، ژانگ س، مک لاوفلین SM، کلمن D، واکر SK، ساتر
CM، et al تعداد تخمک و کمبود تغذیه در محیط اطراف
گوسفند اثرات متفاوتی بر اپنجوتایپ آدرنال، رشد، و
توسعه. Am J Physiol Endocrinol Metab. 2014؛ 307: E141 50.
89 ژانگ S، Rattanatray L، موریسون JL، نیکلاس LM، دروغ S، McMillen IC.
چاقی مادر و علل اولیه چاقی کودکان: وزن کردن
مزایا و هزینه های از دست دادن وزن مادران در periconceptional
دوره برای فرزندان انعطاف پذیری دیابت 2011؛ 2011: 585749.
90 ژانگ S، ویلیامز-ویس O، مک لافلین SM، واکر SK، Kleemann DO، ساتر
CM، et al ناتوانی مادر در طی هفته اول پس از تخمک گذاری
نتیجه گیری در کاهش بیان mRNA گیرنده گلوکوکورتیکوئیدی در
غربالگری hypermethylation اگزون 17 در هیپوفیز جنین در اواخر
حاملگی J Dev Orig Heal Dis. 2013؛ 4:391 401.
91 دروغ S، موریسون JL، Williams-Wyss O، Suter CM، Humphreys DT، Ozanne SE،
و همکاران برنامه های ناباروری Periconceptional تغییر در سیگنالینگ انسولین
مولکول ها و میکرو RNA ها در عضله اسکلتی در تک و دوقلو جنین
گوسفند Biol Reprod. 2014؛ 90: 5.
92 ون استرن EM، ون مایر H، Huijkman NC، ون Dijk TH، Baller JF، Verkade
HJ و همکاران فعال سازی گیرنده های کبد جنین حاد باعث ایجاد لیپوژنز می شود
پاسخ لیپید پلاسمی به رژیم غذایی با چربی بالا در موش بالغ تاثیر نمی گذارد. ام جی
فیزیول اندوکرینول متاب. 2009؛ 297: E1171~8.
93 فرناندز دبلیو دبلیو دبلیو ای، الفارادی MZ، مارتین گرونتر MS، دوکو گیمارس
DE، Piekarz A، Ferland-McCollough D، et al. سرریز IRS-1 در
بافت چربی فرزندان موش های چاق به طور تصادفی برنامه ریزی شده است
از طریق مکانیزم های پست رونویسی مول متاب.
2014 ؛ 3: 325 33.
94 Waterland RA، Travisano M، Tahiliani KG. hypermethylation ناشی از رژیم غذایی در
Agouti زرد زنده قابل انتقال از طریق زن از طریق transgenerationally نیست.
FASEB J. 2007؛ 21:3380~5.
95 ژئو ZJ، لو SM، لین F، لیانگ QX، هوانگ L، Wei YC، و همکاران. متیلاسیون DNA در
تخمک ها و کبد موش های ماده و فرزندان آنها: اثرات رژیم غذایی با چربی بالا
چاقی Env Heal Perspect. 2014؛ 122:159 64.
96. Ollikainen M, Ismail K, Gervin K, Kyllânen A, Hakkarainen A, Lundbom J, et al.
تغییرات متیلاسیون دی ان ای خون در ژنوم در عناصر نظارتی
و مناطق heterochromatic در دوقلوهای monozygotic متضاد برای چاقی
و چربی کبد. اپی ژنتیک کلین 2015 ؛ 7: 1 13.
IFM's Find A Practitioner بزرگترین شبکه ارجاع در پزشکی کاربردی است که برای کمک به بیماران در یافتن پزشکان طب عملکردی در هر نقطه از جهان ایجاد شده است. با توجه به تحصیلات گسترده در پزشکی کاربردی ، پزشکان مجاز IFM اولین بار در نتایج جستجو ذکر شده اند







 چاقی یک بیماری پیچیده و چندوجهی است و درک بهتر مکانیسم های درونی متابولیسم زندگی، محیط زیست و ژنتیک برای ایجاد راهکارهای موثر برای پیشگیری و درمان [1] ضروری است.
چاقی یک بیماری پیچیده و چندوجهی است و درک بهتر مکانیسم های درونی متابولیسم زندگی، محیط زیست و ژنتیک برای ایجاد راهکارهای موثر برای پیشگیری و درمان [1] ضروری است. مدلهای حیوانی فرصتهای منحصر به فردی را برای مطالعات بسیار کنترلشده فراهم میکنند که بینش مکانیکی در مورد نقش علائم اپی ژنتیک خاص، هم به عنوان شاخصهای وضعیت متابولیک فعلی و هم بهعنوان پیشبینیکننده خطر آتی چاقی و بیماریهای متابولیک ارائه میکنند. یکی از جنبههای مهم مطالعات حیوانی این است که آنها امکان ارزیابی تغییرات اپی ژنتیکی در بافتهای هدف، از جمله کبد و هیپوتالاموس را فراهم میکنند، که در انسان بسیار دشوارتر است. علاوه بر این، توانایی برداشت مقادیر زیادی از بافت تازه، ارزیابی چندین علامت کروماتین و همچنین متیلاسیون DNA را ممکن میسازد. برخی از این تغییرات اپی ژنتیکی به تنهایی یا به صورت ترکیبی ممکن است به برنامه ریزی محیطی پاسخ دهند. در مدلهای حیوانی، میتوان چندین نسل از فرزندان را مطالعه کرد و بنابراین تمایز بین انتقال بین نسلی و بین نسلی خطر چاقی را با واسطه حافظه اپی ژنتیکی وضعیت تغذیه والدین، که در مطالعات انسانی به راحتی قابل تشخیص نیست، امکانپذیر کرد. ما از اصطلاح اول برای انتقال میوز خطر در غیاب مواجهه مداوم استفاده می کنیم در حالی که دومی در درجه اول مستلزم انتقال مستقیم خطر از طریق برنامه ریزی مجدد متابولیک جنین یا گامت است.
مدلهای حیوانی فرصتهای منحصر به فردی را برای مطالعات بسیار کنترلشده فراهم میکنند که بینش مکانیکی در مورد نقش علائم اپی ژنتیک خاص، هم به عنوان شاخصهای وضعیت متابولیک فعلی و هم بهعنوان پیشبینیکننده خطر آتی چاقی و بیماریهای متابولیک ارائه میکنند. یکی از جنبههای مهم مطالعات حیوانی این است که آنها امکان ارزیابی تغییرات اپی ژنتیکی در بافتهای هدف، از جمله کبد و هیپوتالاموس را فراهم میکنند، که در انسان بسیار دشوارتر است. علاوه بر این، توانایی برداشت مقادیر زیادی از بافت تازه، ارزیابی چندین علامت کروماتین و همچنین متیلاسیون DNA را ممکن میسازد. برخی از این تغییرات اپی ژنتیکی به تنهایی یا به صورت ترکیبی ممکن است به برنامه ریزی محیطی پاسخ دهند. در مدلهای حیوانی، میتوان چندین نسل از فرزندان را مطالعه کرد و بنابراین تمایز بین انتقال بین نسلی و بین نسلی خطر چاقی را با واسطه حافظه اپی ژنتیکی وضعیت تغذیه والدین، که در مطالعات انسانی به راحتی قابل تشخیص نیست، امکانپذیر کرد. ما از اصطلاح اول برای انتقال میوز خطر در غیاب مواجهه مداوم استفاده می کنیم در حالی که دومی در درجه اول مستلزم انتقال مستقیم خطر از طریق برنامه ریزی مجدد متابولیک جنین یا گامت است.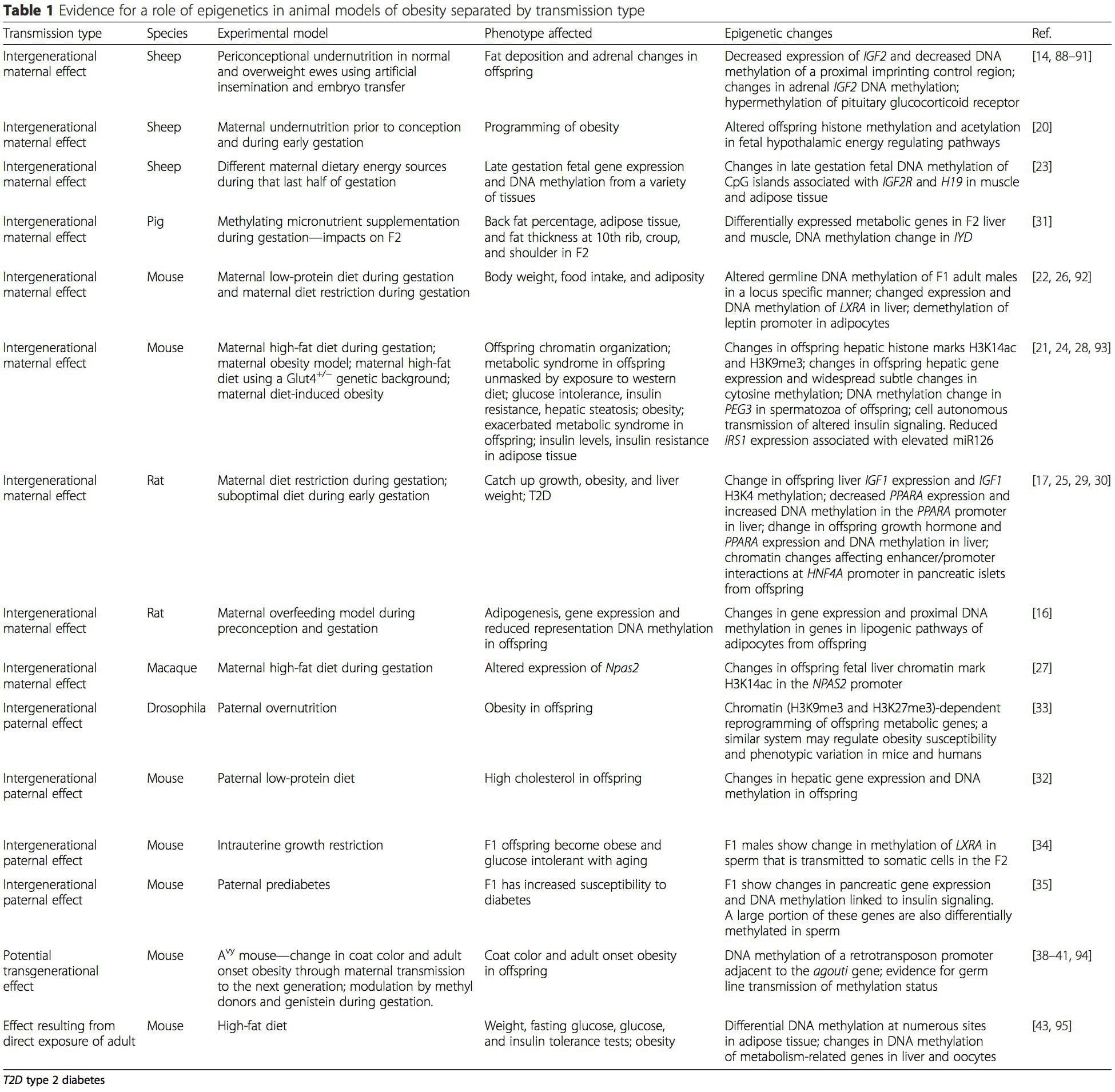 (I) تغييرات اپيگنتيک در پسران همراه با تغذيه مادر در حين بارداري
(I) تغييرات اپيگنتيک در پسران همراه با تغذيه مادر در حين بارداري مکمل های غذایی مادر، سوء تغذیه و تغذیه بیش از حد در دوران بارداری می تواند رسوب چربی و هموستاز انرژی را در فرزندان تغییر دهد [11، 13، 15، 19]. با این اثرات در فرزندان، تغییرات در متیلاسیون DNA، تغییرات پس از ترجمه هیستون، و بیان ژن برای چندین ژن هدف، به ویژه ژن های تنظیم کننده متابولیسم اسیدهای چرب و سیگنال دهی انسولین مرتبط است [16، 17، 20-30]. تنوع مدلهای حیوانی مورد استفاده در این مطالعات و مسیرهای متابولیکی رایج تحت تأثیر قرار گرفته، نشاندهنده یک پاسخ تطبیقی حفظشده تکاملی است که با اصلاح اپی ژنتیکی انجام میشود. با این حال، تعداد کمی از ژنهای شناسایی شده خاص و تغییرات اپی ژنتیکی در مطالعات مرتبط تایید شدهاند، و تحقیقات گسترده در مقیاس ژنوم معمولاً اعمال نشدهاند. یک مانع عمده برای مقایسه این مطالعات، پنجرههای ذهنی مختلف است که در معرض چالشهای تغذیهای قرار میگیرند، که ممکن است نتایج بسیار متفاوتی را ایجاد کند. اثبات اینکه تغییرات اپی ژنتیکی به جای اینکه با تغییرات فنوتیپی فرزندان مرتبط باشد، علت و معلول هستند نیز مورد نیاز است. این امر مستلزم شناسایی پاسخ حافظه اپی ژنتیکی ناشی از تغذیه والدین است که مقدم بر توسعه فنوتیپ تغییر یافته در فرزندان است.
مکمل های غذایی مادر، سوء تغذیه و تغذیه بیش از حد در دوران بارداری می تواند رسوب چربی و هموستاز انرژی را در فرزندان تغییر دهد [11، 13، 15، 19]. با این اثرات در فرزندان، تغییرات در متیلاسیون DNA، تغییرات پس از ترجمه هیستون، و بیان ژن برای چندین ژن هدف، به ویژه ژن های تنظیم کننده متابولیسم اسیدهای چرب و سیگنال دهی انسولین مرتبط است [16، 17، 20-30]. تنوع مدلهای حیوانی مورد استفاده در این مطالعات و مسیرهای متابولیکی رایج تحت تأثیر قرار گرفته، نشاندهنده یک پاسخ تطبیقی حفظشده تکاملی است که با اصلاح اپی ژنتیکی انجام میشود. با این حال، تعداد کمی از ژنهای شناسایی شده خاص و تغییرات اپی ژنتیکی در مطالعات مرتبط تایید شدهاند، و تحقیقات گسترده در مقیاس ژنوم معمولاً اعمال نشدهاند. یک مانع عمده برای مقایسه این مطالعات، پنجرههای ذهنی مختلف است که در معرض چالشهای تغذیهای قرار میگیرند، که ممکن است نتایج بسیار متفاوتی را ایجاد کند. اثبات اینکه تغییرات اپی ژنتیکی به جای اینکه با تغییرات فنوتیپی فرزندان مرتبط باشد، علت و معلول هستند نیز مورد نیاز است. این امر مستلزم شناسایی پاسخ حافظه اپی ژنتیکی ناشی از تغذیه والدین است که مقدم بر توسعه فنوتیپ تغییر یافته در فرزندان است. مطالعات در حال ظهور نشان داده اند که سطح تغذیه پدرانه می تواند بر رسوب چربی و علائم اپی ژنتیک فرزندان تأثیر بگذارد [31-34]. یک تحقیق اخیر با استفاده از موش نشان داده است که پیش دیابت پدرانه منجر به افزایش حساسیت به دیابت در فرزندان F1 با تغییرات مرتبط در بیان ژن لوزالمعده و متیلاسیون DNA مرتبط با سیگنالینگ انسولین می شود [35]. از همه مهمتر ، همپوشانی این تغییرات اپی ژنتیکی در جزایر پانکراس و اسپرم وجود دارد که نشان دهنده وراثت خط میکروب است. با این حال ، بیشتر این مطالعات ، اگرچه از نظر مفاهیم جذاب هستند ، اما در مقیاس ژنومی تحقیق محدود هستند و اغلب تغییرات اپی ژنتیکی ضعیف و تا حدی گذرا را در ارتباط با فنوتیپ های متابولیک خفیف در فرزندان نشان می دهند.
مطالعات در حال ظهور نشان داده اند که سطح تغذیه پدرانه می تواند بر رسوب چربی و علائم اپی ژنتیک فرزندان تأثیر بگذارد [31-34]. یک تحقیق اخیر با استفاده از موش نشان داده است که پیش دیابت پدرانه منجر به افزایش حساسیت به دیابت در فرزندان F1 با تغییرات مرتبط در بیان ژن لوزالمعده و متیلاسیون DNA مرتبط با سیگنالینگ انسولین می شود [35]. از همه مهمتر ، همپوشانی این تغییرات اپی ژنتیکی در جزایر پانکراس و اسپرم وجود دارد که نشان دهنده وراثت خط میکروب است. با این حال ، بیشتر این مطالعات ، اگرچه از نظر مفاهیم جذاب هستند ، اما در مقیاس ژنومی تحقیق محدود هستند و اغلب تغییرات اپی ژنتیکی ضعیف و تا حدی گذرا را در ارتباط با فنوتیپ های متابولیک خفیف در فرزندان نشان می دهند.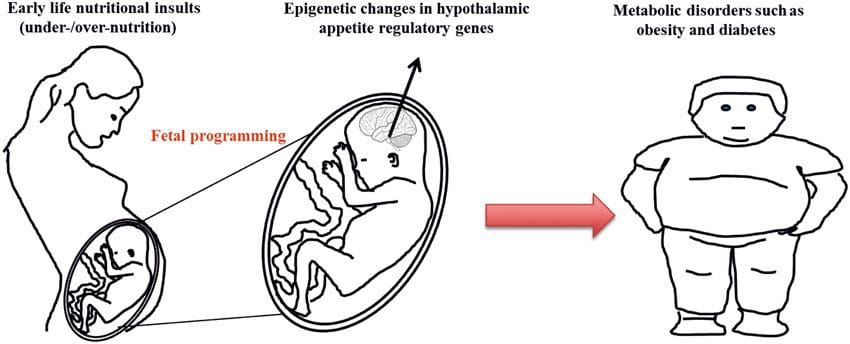 انتقال پایدار اطلاعات اپی ژنتیک در نسل های چندگانه به خوبی در سیستم های گیاهی و C. elegans، اما اهمیت آن در پستانداران هنوز بسیار مورد بحث است [36، 37]. یک مبنای اپی ژنتیکی برای انتقال فنوتیپ ها از پدربزرگ و مادربزرگ در پاسخ به مواجهه با رژیم غذایی به خوبی ایجاد شده است، از جمله در گونه های دام [31]. تأثیرگذارترین مطالعاتی که اثرات انتقال اپی ژنتیکی را بر فنوتیپ فرزندان نشان میدهد، از نمونه موش آگوتی زرد زنده (Avy) استفاده کردهاند [38]. در این موش، قرار دادن یک رتروترانسپوزون در بالادست ژن آگوتی باعث بیان سازنده آن و در نتیجه رنگ زرد پوست و شروع چاقی بزرگسالان می شود. انتقال مادر از طریق خط زایا منجر به خاموش شدن بیان آگوتی به واسطه متیلاسیون DNA می شود که منجر به رنگ پوشش نوع وحشی و فنوتیپ لاغر فرزندان می شود [39، 40]. نکته مهم، مطالعات بعدی در این موش ها نشان داد که قرار گرفتن مادر در معرض اهداکنندگان متیل باعث تغییر رنگ پوشش می شود [41]. یک مطالعه انتقال یک فنوتیپ به نسل F3 و تغییرات در بیان تعداد زیادی ژن در پاسخ به محدودیت پروتئین در F0 را گزارش کرده است [42]. با این حال، تغییرات در بیان بسیار متغیر بود و ارتباط مستقیمی با تغییرات اپی ژنتیکی در این سیستم شناسایی نشد.
انتقال پایدار اطلاعات اپی ژنتیک در نسل های چندگانه به خوبی در سیستم های گیاهی و C. elegans، اما اهمیت آن در پستانداران هنوز بسیار مورد بحث است [36، 37]. یک مبنای اپی ژنتیکی برای انتقال فنوتیپ ها از پدربزرگ و مادربزرگ در پاسخ به مواجهه با رژیم غذایی به خوبی ایجاد شده است، از جمله در گونه های دام [31]. تأثیرگذارترین مطالعاتی که اثرات انتقال اپی ژنتیکی را بر فنوتیپ فرزندان نشان میدهد، از نمونه موش آگوتی زرد زنده (Avy) استفاده کردهاند [38]. در این موش، قرار دادن یک رتروترانسپوزون در بالادست ژن آگوتی باعث بیان سازنده آن و در نتیجه رنگ زرد پوست و شروع چاقی بزرگسالان می شود. انتقال مادر از طریق خط زایا منجر به خاموش شدن بیان آگوتی به واسطه متیلاسیون DNA می شود که منجر به رنگ پوشش نوع وحشی و فنوتیپ لاغر فرزندان می شود [39، 40]. نکته مهم، مطالعات بعدی در این موش ها نشان داد که قرار گرفتن مادر در معرض اهداکنندگان متیل باعث تغییر رنگ پوشش می شود [41]. یک مطالعه انتقال یک فنوتیپ به نسل F3 و تغییرات در بیان تعداد زیادی ژن در پاسخ به محدودیت پروتئین در F0 را گزارش کرده است [42]. با این حال، تغییرات در بیان بسیار متغیر بود و ارتباط مستقیمی با تغییرات اپی ژنتیکی در این سیستم شناسایی نشد.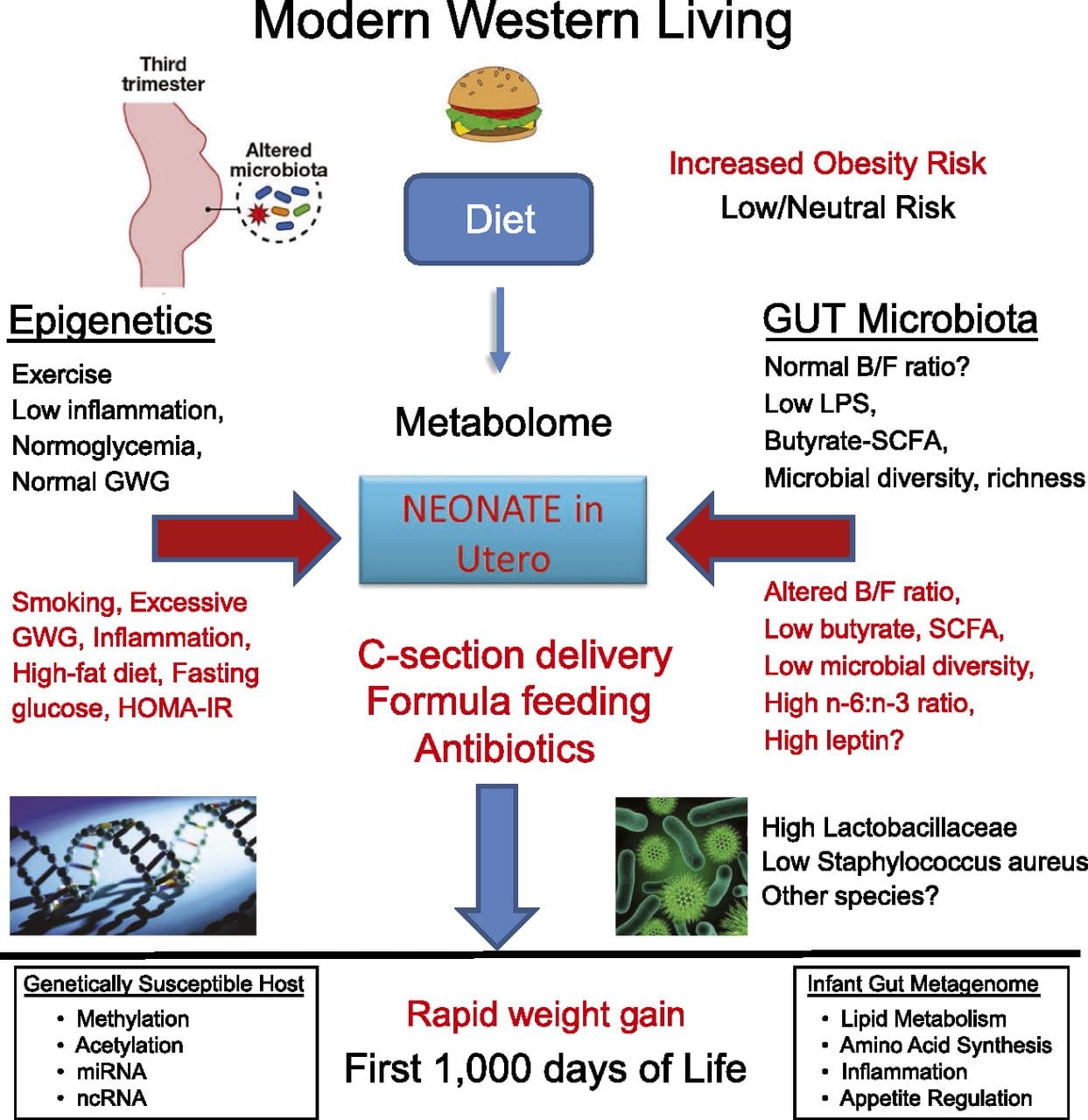 در حالی که بسیاری از مطالعات تغییرات اپی ژنتیکی مرتبط با رژیم را در مدل های حیوانی با استفاده از مناطق خاص منطقه نامشخص نشان داده اند، تجزیه و تحلیل چندین ژنوم انجام شده است. مطالعات اخیر بر روی تعیین اثر مستقیم اپی ژنتیکی رژیم های غذایی با چربی بالا / چاقی ناشی از رژیم غذایی در موش های بالغ با استفاده از بیان ژن ژنوم و تجزیه و تحلیل متیلاسیون DNA [43] متمرکز شده است. در این مطالعه، 232 به طور متفاوتی متوله شده (DMRs) در آديپوسیت ها از موش های کنترل و پرورش دهنده با چربی بالا شناخته شد. مهمتر از آن، مناطق مربوطه انسان برای DMR های موش نیز در بافت چربی از جمعیت چاق و لاغر متhylly متhylized شده، در نتیجه برجسته حفاظت تکاملی قابل توجه این مناطق. این نتیجه بر اهمیت احتمال DMR های شناسایی شده در تنظیم هومیوستاز انرژی در پستانداران تاکید دارد.
در حالی که بسیاری از مطالعات تغییرات اپی ژنتیکی مرتبط با رژیم را در مدل های حیوانی با استفاده از مناطق خاص منطقه نامشخص نشان داده اند، تجزیه و تحلیل چندین ژنوم انجام شده است. مطالعات اخیر بر روی تعیین اثر مستقیم اپی ژنتیکی رژیم های غذایی با چربی بالا / چاقی ناشی از رژیم غذایی در موش های بالغ با استفاده از بیان ژن ژنوم و تجزیه و تحلیل متیلاسیون DNA [43] متمرکز شده است. در این مطالعه، 232 به طور متفاوتی متوله شده (DMRs) در آديپوسیت ها از موش های کنترل و پرورش دهنده با چربی بالا شناخته شد. مهمتر از آن، مناطق مربوطه انسان برای DMR های موش نیز در بافت چربی از جمعیت چاق و لاغر متhylly متhylized شده، در نتیجه برجسته حفاظت تکاملی قابل توجه این مناطق. این نتیجه بر اهمیت احتمال DMR های شناسایی شده در تنظیم هومیوستاز انرژی در پستانداران تاکید دارد.
 (i) مطالعات ارتباط ژنتیکی. پلیمورفیسمهای ژنتیکی که با افزایش خطر ابتلا به بیماریهای خاص مرتبط هستند، به طور پیشینی با ژنهای عامل مرتبط هستند. وجود متیلاسیون افتراقی در چنین مناطقی ارتباط عملکردی این تغییرات اپی ژنتیکی را در کنترل بیان ژن(های پروگزیمال) استنباط می کند. اثرات ژنتیکی قوی با اثر سیس وجود دارد که زیربنای بسیاری از تنوع اپی ژنتیکی است [7، 45]، و در مطالعات مبتنی بر جمعیت، روشهایی که از جانشینهای ژنتیکی برای استنباط نقش علّی یا واسطهای تفاوتهای اپی ژنوم استفاده میکنند [7، 46-48] استفاده شده است. . استفاده از اطلاعات ژنتیکی خانوادگی همچنین میتواند منجر به شناسایی مناطق کاندید بالقوه مسبب شود که متیلاسیون دیفرانسیل مرتبط با فنوتیپ را نشان میدهند [49].
(i) مطالعات ارتباط ژنتیکی. پلیمورفیسمهای ژنتیکی که با افزایش خطر ابتلا به بیماریهای خاص مرتبط هستند، به طور پیشینی با ژنهای عامل مرتبط هستند. وجود متیلاسیون افتراقی در چنین مناطقی ارتباط عملکردی این تغییرات اپی ژنتیکی را در کنترل بیان ژن(های پروگزیمال) استنباط می کند. اثرات ژنتیکی قوی با اثر سیس وجود دارد که زیربنای بسیاری از تنوع اپی ژنتیکی است [7، 45]، و در مطالعات مبتنی بر جمعیت، روشهایی که از جانشینهای ژنتیکی برای استنباط نقش علّی یا واسطهای تفاوتهای اپی ژنوم استفاده میکنند [7، 46-48] استفاده شده است. . استفاده از اطلاعات ژنتیکی خانوادگی همچنین میتواند منجر به شناسایی مناطق کاندید بالقوه مسبب شود که متیلاسیون دیفرانسیل مرتبط با فنوتیپ را نشان میدهند [49].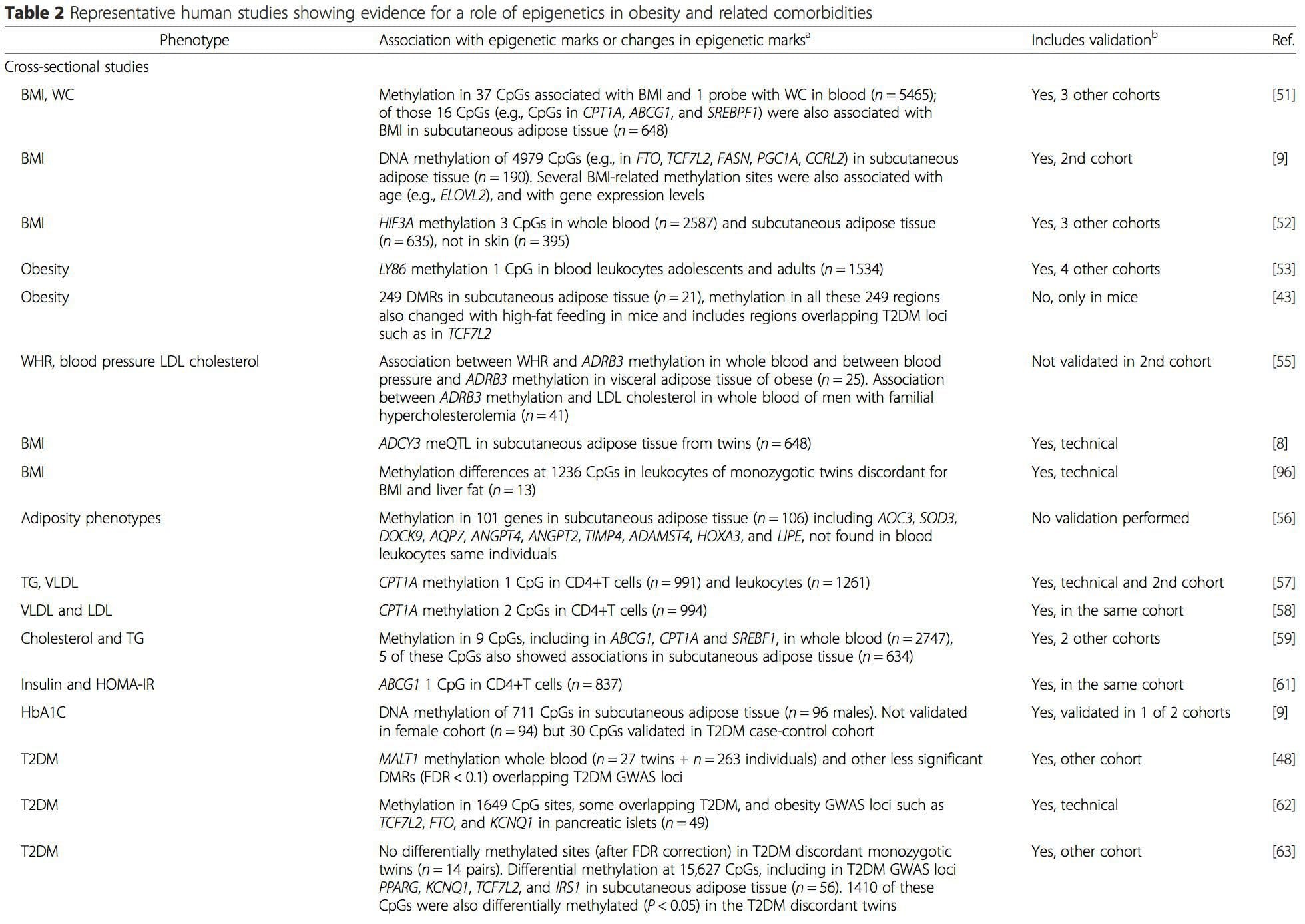
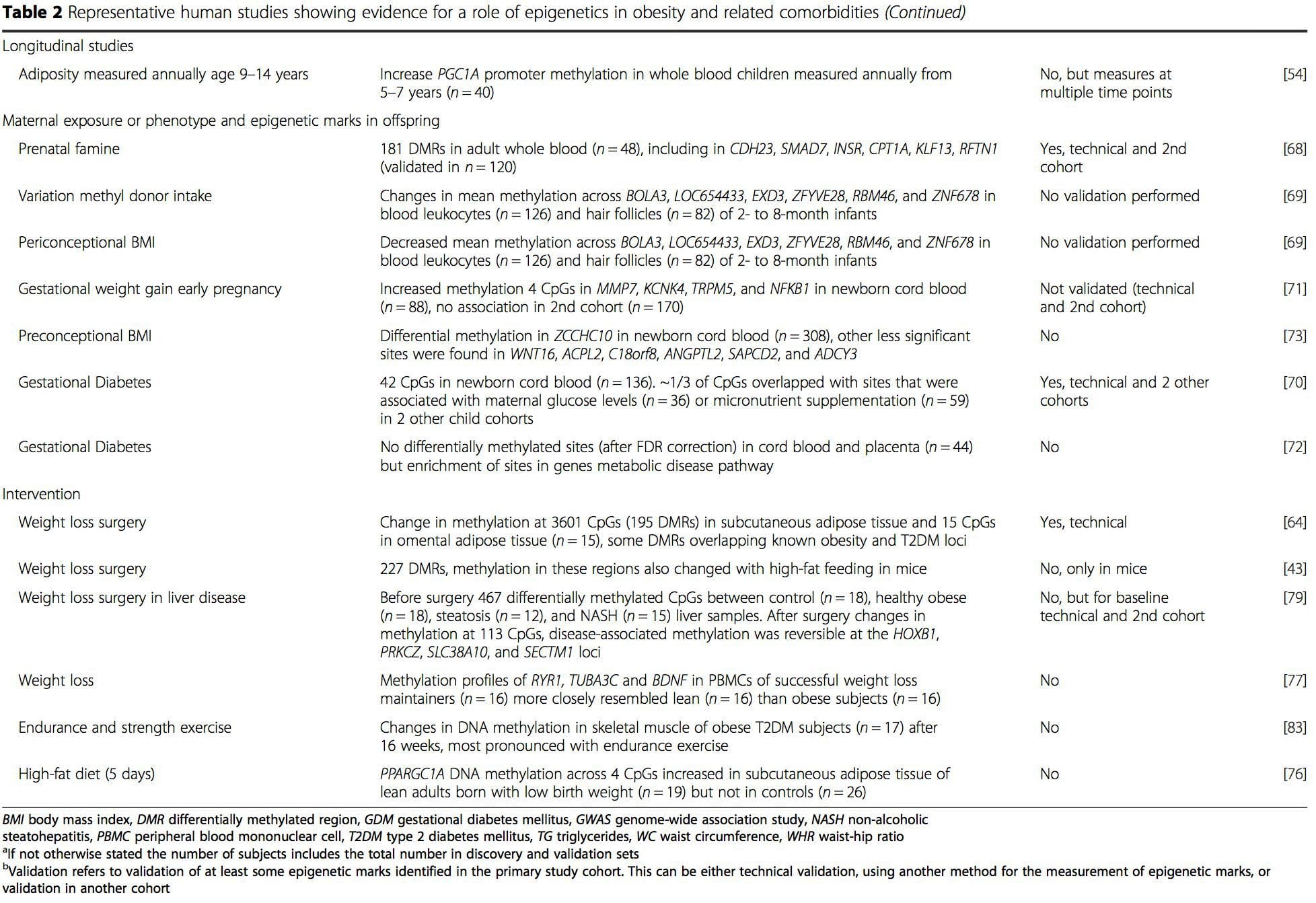 از این مطالعات، متیلاسیون تغییرات PGC1A، HIF3A، ABCG1 و CPT1A و RXRA [18] که قبلا توصیف شده است، به عنوان نشانگرهای زیستی مرتبط با سلامت متابولیک یا شاید پیش بینی کننده آن، متاثر از کاندیداهای قابل قبول برای نقش در توسعه بیماری متابولیک .
از این مطالعات، متیلاسیون تغییرات PGC1A، HIF3A، ABCG1 و CPT1A و RXRA [18] که قبلا توصیف شده است، به عنوان نشانگرهای زیستی مرتبط با سلامت متابولیک یا شاید پیش بینی کننده آن، متاثر از کاندیداهای قابل قبول برای نقش در توسعه بیماری متابولیک .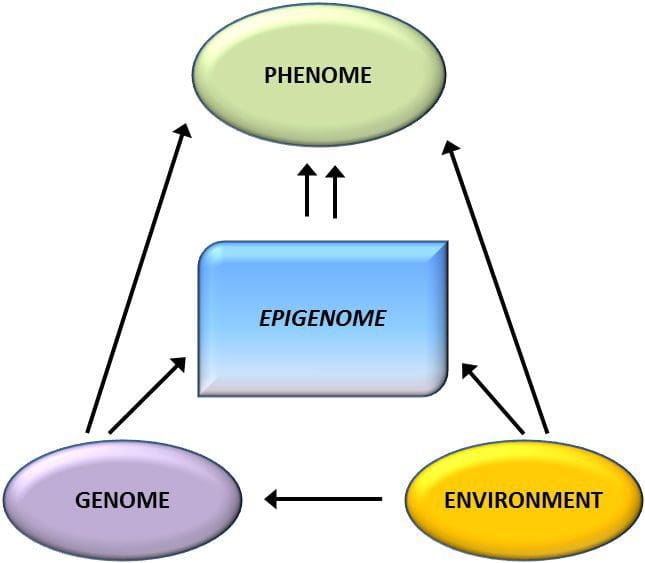 تنوع اپی ژنتیکی به شدت تحتتاثیر تنوع ژنتیکی زیربنایی قرار میگیرد، با تخمین زده میشود که ژنوتیپ 20 تا 40 درصد از تغییرات را توضیح دهد [6، 8]. اخیراً، تعدادی از مطالعات شروع به ادغام دادههای متیلوم و ژنوتیپ برای شناسایی مکانهای صفت کمی متیلاسیون (meQTL) مرتبط با فنوتیپهای بیماری کردهاند. به عنوان مثال، در بافت چربی، یک meQTL همپوشانی با منبع خطر ژنتیکی BMI در یک عنصر تقویت کننده بالادست ADCY3 شناسایی شده است [8]. مطالعات دیگر همچنین همپوشانی هایی را بین چاقی شناخته شده و جایگاه های خطر T2DM و DMR های مرتبط با چاقی و T2DM شناسایی کرده اند [43، 48، 62]. متیلاسیون تعدادی از این DMRها نیز با تغذیه با چربی بالا در موش [43] و کاهش وزن در انسان [64] تعدیل شد. این نتایج یک پیوند جالب بین تغییرات ژنتیکی مرتبط با حساسیت به بیماری و ارتباط آنها با مناطقی از ژنوم را که در پاسخ به چالشهای تغذیهای دستخوش تغییرات اپی ژنتیکی میشوند، مشخص میکند که دلالت بر یک رابطه علّی دارد. ارتباط نزدیک بین تنوع ژنتیکی و اپی ژنتیکی ممکن است نشان دهنده نقش اساسی آنها در ایجاد تنوع فردی باشد [65، 66]. با این حال، در حالی که این یافته ها نشان می دهد که متیلاسیون DNA ممکن است واسطه اثرات ژنتیکی باشد، همچنین مهم است که در نظر بگیریم که هر دو فرآیند ژنتیکی و اپی ژنتیکی می توانند به طور مستقل بر روی ژن های یکسان عمل کنند. مطالعات دوقلو [8، 63، 67] میتوانند بینشهای مهمی ارائه دهند و نشان دهند که تفاوتهای بین فردی در سطوح متیلاسیون DNA عمدتاً از محیط غیرمشترک و تأثیرات تصادفی ناشی میشود، حداقل از اثرات محیطی مشترک، اما همچنین با تأثیر قابل توجه ژنتیکی. تغییر.
تنوع اپی ژنتیکی به شدت تحتتاثیر تنوع ژنتیکی زیربنایی قرار میگیرد، با تخمین زده میشود که ژنوتیپ 20 تا 40 درصد از تغییرات را توضیح دهد [6، 8]. اخیراً، تعدادی از مطالعات شروع به ادغام دادههای متیلوم و ژنوتیپ برای شناسایی مکانهای صفت کمی متیلاسیون (meQTL) مرتبط با فنوتیپهای بیماری کردهاند. به عنوان مثال، در بافت چربی، یک meQTL همپوشانی با منبع خطر ژنتیکی BMI در یک عنصر تقویت کننده بالادست ADCY3 شناسایی شده است [8]. مطالعات دیگر همچنین همپوشانی هایی را بین چاقی شناخته شده و جایگاه های خطر T2DM و DMR های مرتبط با چاقی و T2DM شناسایی کرده اند [43، 48، 62]. متیلاسیون تعدادی از این DMRها نیز با تغذیه با چربی بالا در موش [43] و کاهش وزن در انسان [64] تعدیل شد. این نتایج یک پیوند جالب بین تغییرات ژنتیکی مرتبط با حساسیت به بیماری و ارتباط آنها با مناطقی از ژنوم را که در پاسخ به چالشهای تغذیهای دستخوش تغییرات اپی ژنتیکی میشوند، مشخص میکند که دلالت بر یک رابطه علّی دارد. ارتباط نزدیک بین تنوع ژنتیکی و اپی ژنتیکی ممکن است نشان دهنده نقش اساسی آنها در ایجاد تنوع فردی باشد [65، 66]. با این حال، در حالی که این یافته ها نشان می دهد که متیلاسیون DNA ممکن است واسطه اثرات ژنتیکی باشد، همچنین مهم است که در نظر بگیریم که هر دو فرآیند ژنتیکی و اپی ژنتیکی می توانند به طور مستقل بر روی ژن های یکسان عمل کنند. مطالعات دوقلو [8، 63، 67] میتوانند بینشهای مهمی ارائه دهند و نشان دهند که تفاوتهای بین فردی در سطوح متیلاسیون DNA عمدتاً از محیط غیرمشترک و تأثیرات تصادفی ناشی میشود، حداقل از اثرات محیطی مشترک، اما همچنین با تأثیر قابل توجه ژنتیکی. تغییر. محیط زیست قبل از زایمان: دو مطالعه که اخیراً منتشر شده است ، از جمعیت انسانی استفاده می کند که تغییرات طبیعی در تأمین مواد مغذی را تجربه کرده اند تا تأثیر تغذیه مادر قبل یا حین بارداری بر متیلاسیون DNA در فرزندان را بررسی کند. اولین مطالعه با استفاده از یک گروه مادر و کودک گامبیایی نشان داد که هر دو تغییر فصلی در مصرف متیل دهنده مادر در دوران بارداری و BMI مادر قبل از بارداری با تغییر متیلاسیون در نوزادان ارتباط دارد [68]. مطالعه دوم با استفاده از فرزندان بزرگسال از گروه گرسنگی گرسنگی هلندی برای بررسی اثر قرار گرفتن در معرض بارداری در یک دوره حاد سو und تغذیه شدید مادر بر متیلاسیون DNA ژن های درگیر در رشد و متابولیسم در بزرگسالی [69]. نتایج اهمیت زمان قرار گرفتن در معرض را در تأثیر آن بر اپی ژنوم برجسته کرد ، زیرا اثرات قابل توجه اپی ژنتیکی فقط در افرادی که در اوایل حاملگی در معرض قحطی قرار داشتند ، مشخص شد. نکته مهم ، تغییرات اپی ژنتیکی همراه با افزایش BMI است. با این حال ، نمی توان در این مطالعه مشخص کرد که آیا این تغییرات در اوایل زندگی وجود داشته اند یا نتیجه یک BMI بالاتر است.
محیط زیست قبل از زایمان: دو مطالعه که اخیراً منتشر شده است ، از جمعیت انسانی استفاده می کند که تغییرات طبیعی در تأمین مواد مغذی را تجربه کرده اند تا تأثیر تغذیه مادر قبل یا حین بارداری بر متیلاسیون DNA در فرزندان را بررسی کند. اولین مطالعه با استفاده از یک گروه مادر و کودک گامبیایی نشان داد که هر دو تغییر فصلی در مصرف متیل دهنده مادر در دوران بارداری و BMI مادر قبل از بارداری با تغییر متیلاسیون در نوزادان ارتباط دارد [68]. مطالعه دوم با استفاده از فرزندان بزرگسال از گروه گرسنگی گرسنگی هلندی برای بررسی اثر قرار گرفتن در معرض بارداری در یک دوره حاد سو und تغذیه شدید مادر بر متیلاسیون DNA ژن های درگیر در رشد و متابولیسم در بزرگسالی [69]. نتایج اهمیت زمان قرار گرفتن در معرض را در تأثیر آن بر اپی ژنوم برجسته کرد ، زیرا اثرات قابل توجه اپی ژنتیکی فقط در افرادی که در اوایل حاملگی در معرض قحطی قرار داشتند ، مشخص شد. نکته مهم ، تغییرات اپی ژنتیکی همراه با افزایش BMI است. با این حال ، نمی توان در این مطالعه مشخص کرد که آیا این تغییرات در اوایل زندگی وجود داشته اند یا نتیجه یک BMI بالاتر است. محیط زیست پس از زایمان: اپی ژنوم در طی رشد جنینی از نو تأسیس می شود و بنابراین ، به احتمال زیاد محیط قبل از تولد بیشترین تأثیر را روی اپی ژنوم دارد. با این حال ، اکنون مشخص شده است که تحت تأثیر طیف وسیعی از شرایط ، از جمله پیری ، قرار گرفتن در معرض سموم و تغییرات رژیم غذایی ، تغییراتی در اپی ژنوم بالغ ایجاد می شود. به عنوان مثال ، تغییرات متیلاسیون DNA در ژنهای متعدد در عضله اسکلتی و PGC1A در بافت چربی در پاسخ به رژیم غذایی پرچرب نشان داده شده است [75 ، 76]. مداخلات برای از دست دادن توده چربی بدن نیز با تغییر در متیلاسیون DNA همراه بوده است. مطالعات گزارش داده اند که پروفایل متیلاسیون DNA بافت چربی [43 ، 64] ، سلولهای تک هسته ای خون محیطی [77] و بافت عضلانی [78] در بیماران چاق قبلاً بیشتر به مشخصات افراد لاغر بدنبال کاهش وزن شباهت دارد. جراحی کاهش وزن همچنین تغییرات متیلاسیون مرتبط با بیماری کبد چرب غیر الکلی را تا حدی معکوس کرد [79] و در مطالعه دیگری منجر به هیپومتیلاسیون ژن های متعدد کاندید چاقی شد ، در مقایسه با چربی مطلق (احشایی) اثرات بارزتری در زیر جلدی داشت [64] . شواهد جمع آوری شده نشان می دهد که مداخلات ورزشی همچنین می تواند بر متیلاسیون DNA تأثیر بگذارد. بیشتر این مطالعات در افراد لاغر انجام شده است [80-82] ، اما یک مطالعه ورزشی در افراد چاق T2DM همچنین تغییراتی در متیلاسیون DNA را نشان داده است ، از جمله در ژن های درگیر در انتقال اسیدهای چرب و گلوکز [83]. تغییرات اپی ژنتیکی نیز با افزایش سن رخ می دهد ، و داده های اخیر نقش چاقی در افزایش آنها را نشان می دهد [9 ، 84 ، 85]. چاقی ، سن اپی ژنتیک بافت کبد را تسریع می کند ، اما بر خلاف یافته های توضیح داده شده در بالا ، این اثر پس از کاهش وزن قابل برگشت نیست [84].
محیط زیست پس از زایمان: اپی ژنوم در طی رشد جنینی از نو تأسیس می شود و بنابراین ، به احتمال زیاد محیط قبل از تولد بیشترین تأثیر را روی اپی ژنوم دارد. با این حال ، اکنون مشخص شده است که تحت تأثیر طیف وسیعی از شرایط ، از جمله پیری ، قرار گرفتن در معرض سموم و تغییرات رژیم غذایی ، تغییراتی در اپی ژنوم بالغ ایجاد می شود. به عنوان مثال ، تغییرات متیلاسیون DNA در ژنهای متعدد در عضله اسکلتی و PGC1A در بافت چربی در پاسخ به رژیم غذایی پرچرب نشان داده شده است [75 ، 76]. مداخلات برای از دست دادن توده چربی بدن نیز با تغییر در متیلاسیون DNA همراه بوده است. مطالعات گزارش داده اند که پروفایل متیلاسیون DNA بافت چربی [43 ، 64] ، سلولهای تک هسته ای خون محیطی [77] و بافت عضلانی [78] در بیماران چاق قبلاً بیشتر به مشخصات افراد لاغر بدنبال کاهش وزن شباهت دارد. جراحی کاهش وزن همچنین تغییرات متیلاسیون مرتبط با بیماری کبد چرب غیر الکلی را تا حدی معکوس کرد [79] و در مطالعه دیگری منجر به هیپومتیلاسیون ژن های متعدد کاندید چاقی شد ، در مقایسه با چربی مطلق (احشایی) اثرات بارزتری در زیر جلدی داشت [64] . شواهد جمع آوری شده نشان می دهد که مداخلات ورزشی همچنین می تواند بر متیلاسیون DNA تأثیر بگذارد. بیشتر این مطالعات در افراد لاغر انجام شده است [80-82] ، اما یک مطالعه ورزشی در افراد چاق T2DM همچنین تغییراتی در متیلاسیون DNA را نشان داده است ، از جمله در ژن های درگیر در انتقال اسیدهای چرب و گلوکز [83]. تغییرات اپی ژنتیکی نیز با افزایش سن رخ می دهد ، و داده های اخیر نقش چاقی در افزایش آنها را نشان می دهد [9 ، 84 ، 85]. چاقی ، سن اپی ژنتیک بافت کبد را تسریع می کند ، اما بر خلاف یافته های توضیح داده شده در بالا ، این اثر پس از کاهش وزن قابل برگشت نیست [84]. تغییرات متیلاسیون DNA در ارتباط با چاقی یا ناشی از رژیم های غذایی یا مداخلات در سبک زندگی و کاهش وزن معمولاً ناچیز است (<15٪) ، اگرچه این بسته به فنوتیپ و بافت مورد مطالعه متفاوت است. به عنوان مثال ، تغییرات بیشتر از 20٪ در بافت چربی پس از کاهش وزن گزارش شده است [64] و ارتباط بین متیلاسیون HIF3A و BMI در بافت چربی بیشتر از خون بود [52].
تغییرات متیلاسیون DNA در ارتباط با چاقی یا ناشی از رژیم های غذایی یا مداخلات در سبک زندگی و کاهش وزن معمولاً ناچیز است (<15٪) ، اگرچه این بسته به فنوتیپ و بافت مورد مطالعه متفاوت است. به عنوان مثال ، تغییرات بیشتر از 20٪ در بافت چربی پس از کاهش وزن گزارش شده است [64] و ارتباط بین متیلاسیون HIF3A و BMI در بافت چربی بیشتر از خون بود [52].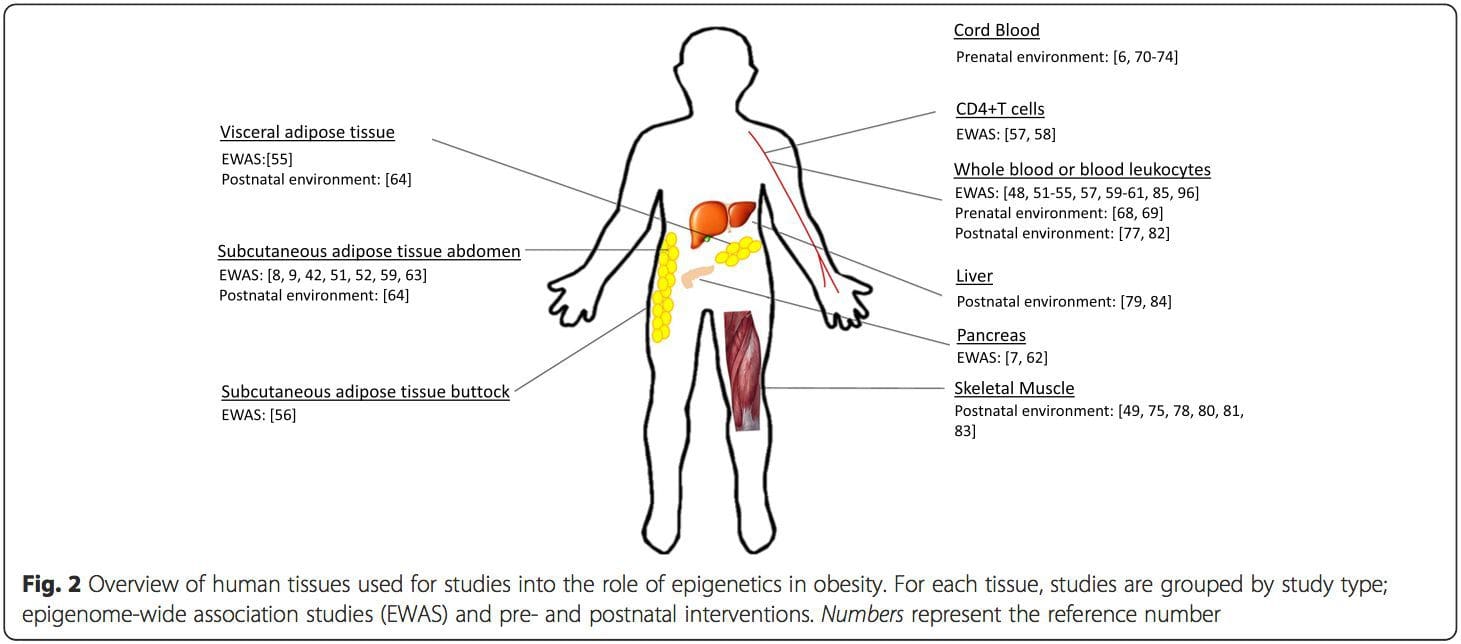 نتیجه گیری
نتیجه گیری




