پشتیبان کلینیک نوروفیزیولوژی بالینی. ال پاسو، TX. دکتر الکساندر جیمنز، معاون درمانگر، بحث می کند نوروفیزیولوژی بالینی دکتر جیمنز اهمیت بالینی و فعالیت های عملکردی فیبرهای عصبی محیطی، نخاعی، مغز و مغز را در زمینه اختلالات عصبی و اسکلتی عضلانی بررسی خواهد کرد. بیماران درک پیشرفته ای از آناتومی، ژنتیک، بیوشیمی و فیزیولوژی درد در ارتباط با سندرم های مختلف بالینی به دست می آورند. بیوشیمی تغذیه ای در ارتباط با غلبه بر درد و درد خواهد شد. و اجرای این اطلاعات به برنامه های درمانی تأکید خواهد شد.
تیم ما از ارائه پروتکل های درمانی اثبات شده به خانواده ها و بیماران مجروح ما بسیار افتخار می کند. با آموزش سلامت کامل به عنوان یک سبک زندگی، ما نه تنها زندگی بیماران، بلکه خانواده های آنها را نیز تغییر می دهیم. ما این کار را انجام می دهیم تا بدون توجه به مسائل مربوط به مقرون به صرفه بودن، به تعداد زیادی از ال پاسوایی هایی که به ما نیاز دارند دسترسی پیدا کنیم. برای پاسخ به هر گونه سؤالی که ممکن است داشته باشید، لطفاً با دکتر جیمنز با شماره 915-850-0900 تماس بگیرید.
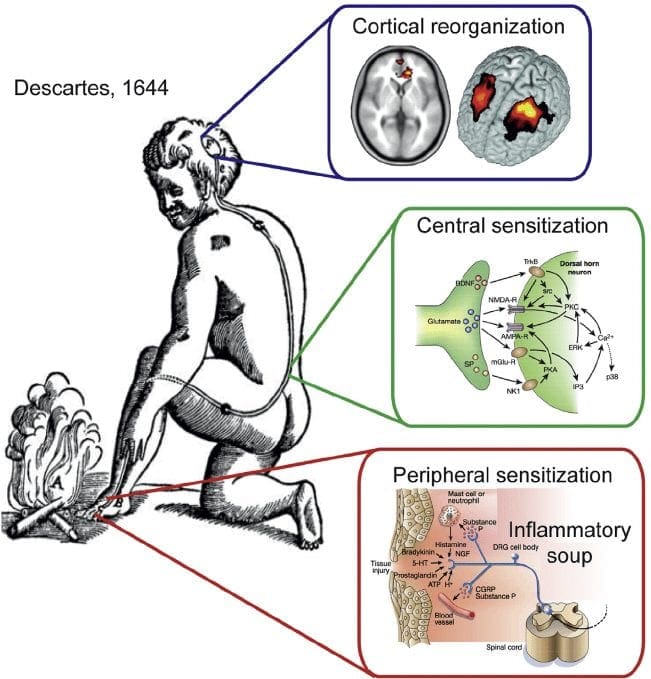
درد نوروپاتیک یک بیماری پیچیده و مزمن درد است که معمولا با آسیب بافت نرم همراه است. درد نوروپاتیک در عمل بالینی رایج است و همچنین برای بیماران و پزشکان به طور یکسان چالش دارد. با درد نوروپاتیک، الیاف عصبی خود می توانند آسیب دیده، ناکارآمد یا مجروح شوند. درد نوروپاتیک نتیجه آسیب های ناشی از تروما یا بیماری به سیستم عصبی محیطی یا محیطی است که در آن ضایعه ممکن است در هر محل رخ دهد. در نتیجه، این فیبرهای عصبی آسیب دیده می توانند سیگنال های نادرست را به مراکز درمانی دیگر ارسال کنند. اثر یک آسیب نخاع عصبی شامل تغییر در عملکرد عصبی، هر دو در منطقه آسیب و همچنین در اطراف آسیب است. علائم بالینی درد نوروپاتیک معمولا شامل پدیده های حسی مانند درد خودبه خود، پاراستزی و هیپرالژزی می شود.
درد نوروپاتیک، به عنوان تعریف شده توسط انجمن بین المللی مطالعه درد یا IASP، درد آغاز شده و یا ناشی از ضایعه اولیه یا اختلال عملکرد سیستم عصبی است. این می تواند از آسیب در هر نقطه در امتداد neuraxis: سیستم عصبی محیطی، سیستم عصبی ستون فقرات و یا supraspinal ایجاد شود. ویژگی هایی که درد نوروپاتیک را از سایر انواع درد ها تشخیص می دهند، شامل درد و علائم حسی است که فراتر از دوره بهبودی است. این در انسان به علت درد خود به خود، آلدوینیا یا تجربه تحریک غیرخوابی به عنوان دردناک و علت درد و یا درد سوزش مداوم مشخص می شود. درد خودبهخودی شامل احساسات "سوزن و سوزن"، سوزش، تیراندازی، پوسیدگی و درد پره اکزمیال یا ضربه الکتریکی مانند درد است که اغلب با دیسستی و پارستزی همراه است. این احساسات نه تنها دستگاه حسی بیمار را تغییر می دهد، بلکه همچنین به سلامت، خلق، توجه و تفکر بیمار نیز می پردازد. درد نوروپاتیک از علائم "منفی" مانند حساسیت و احساس سوزن شدن و علائم "مثبت" مانند پارستزی، درد خودبه خودی و افزایش درد احساس می شود.
شرایطی که غالبا مربوط به درد نوروپاتیک است می توان به دو گروه اصلی تقسیم کرد: درد ناشی از آسیب سیستم عصبی مرکزی و درد به علت آسیب به سیستم عصبی محیطی. سکته های مغزی و زیر قشر، آسیب های آسیب دیده نخاعی، سیرینگو میلایا و سیرینگووبولبیا، نورالژی های سه گانه و گلوزوفارنکس، ضایعات نئوپلاستی و سایر اشعه های فضایی، شرایط بالینی است که متعلق به گروه سابق است. نوروپاتی عصبی، نروپاتی، ناهنجاریهای عصبی، پلیمریپاتیهای محیطی، پلاكوپاتی، فشرده سازی ریشه عصب، پارگی بعد از آمپوتاسیون و درد اندام فانتوم، ناخوشی پس از فروپاشیدن و نوروپاتیهای مرتبط با سرطان، شرایط بالینی متعلق به گروه دوم هستند.
پاتوفیزیولوژی درد نوروپاتیک
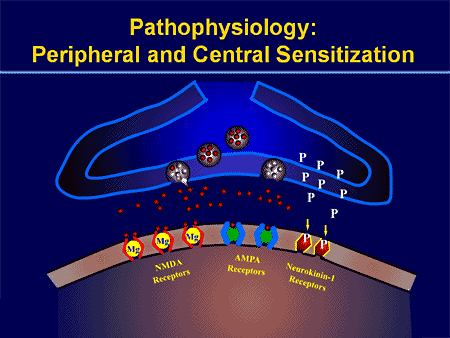
فرآیندهای پاتوفیزیولوژیک و مفاهیم مبتنی بر درد نوروپاتیک چندگانه هستند. قبل از پوشش دادن این فرایندها، بازبینی مدارهای معمولی درد مهم است. مدارهای منظم درد عبارتند از فعال سازی یک عفونت بدون علامت، که همچنین به عنوان گیرنده درد شناخته می شود، در پاسخ به تحریک دردناک. موج دپولاریزاسیون به نورونهای اولویتی تحویل داده می شود، همراه با سدیم بوسیله کانال های سدیم و عجله پتاسیم. نورونها در مغز مغز در هسته سه گانه یا در شاخ پشتی نخاع به پایان می رسند. این در اینجا جایی است که نشانه کانال های کلسیم ولتاژ را در ترمینال پیش سیناپتیک باز می کند، که اجازه می دهد کلسیم وارد شود. کلسیم اجازه می دهد گلوتامات، انتقال دهنده عصبی هیجان انگیز، به منطقه سیناپسی آزاد شود. گلوتامات به گیرنده های NMDA در نورون های دوم مرتبه اتصال می دهد و باعث ایجاد ضریب تخمیر می شود.
این نورون ها از طناب نخاعی عبور می کنند و تا تالاموس حرکت می کنند، جایی که آنها با نورون های ثانویه سیناپس هماهنگ می شوند. اینها به سیستم لیمبیک و قشر مغزی متصل می شوند. همچنین یک مسیر مهاری وجود دارد که مانع انتقال سیگنال درد از شاخ پشتی می شود. نورونهای ضد حساسیت از مغز مغز استخراج میشوند و با استفاده از دوپامین و نوراپی نفرین، کمر نخاعی را در جایی که با اینترنوریونهای کوتاه در شاخ پشتی سیناپس می کنند، حرکت می کنند. اینترنئورون ها سیناپس بین نورون اول و همچنین نورون دوم مرتبه را با انتشار اسید گاما آمینو بوتیریک یا GABA، انتقال دهنده عصبی مهار می کنند. درنتیجه، توقف درد ناشی از مهار سیناپس ها بین نورون های اول و دوم می باشد، در حالیکه افزایش درد ممکن است در نتیجه سرکوب اتصالات مهار کننده سیناپسی باشد.
با این حال، مکانیسم درد نوروپاتیک، روشن نیست. مطالعات متعدد حیوانی نشان داده است که بسیاری از مکانیسم ها ممکن است درگیر شوند با این حال، باید به یاد داشته باشید که آنچه که در مورد موجودات وجود دارد، همیشه برای مردم اعمال نمی شود. نورونهای مرتبه اول ممکن است شلیک خود را افزایش دهند اگر آنها تا حدی آسیب دیده و میزان کانال های سدیم را افزایش دهند. تخلیه خارج از رحم ناشی از افزایش دپولاریزاسیون در محل های خاصی در فیبر است که موجب درد و خود درد و درد ناشی از حرکت می شود. مدارهای ممنوعه ممکن است در سطح شاخ پشتی یا سلول های بنیادی مغز کاهش یابد، و همچنین هر دو، اجازه می دهد که درد های ناخواسته به سفر بیفتند.
علاوه بر این، در پردازش مرکزی درد ممکن است تغییر کند، در حالی که به علت درد مزمن و استفاده از برخی داروها و یا داروها، نورون های ثانویه و سوم می توانند "حافظه" درد و حساسیت ایجاد کنند. سپس حساسیت شدید نورون های نخاعی و آستانه فعال سازی کاهش می یابد. نظریه دیگری نشان می دهد که مفهوم درد نوروپاتیک با همدردی حفظ شده است. این مفهوم با درد زدائی پس از سمپاتکتومی از حیوانات و مردم نشان داده شد. با این حال، ترکیبی از مکانیک می تواند در بسیاری از درد مزمن نوروپاتیک یا مخلوط سنتی و نوروپاتیک درگیر شود. از جمله چالش های موجود در زمینه درد و خیلی بیشتر آنکه به درد نوروپاتیک مربوط می شود، توانایی آن است که آن را بررسی کنید. برای این دو جزء وجود دارد: اول، ارزیابی کیفیت، شدت و پیشرفت؛ و دوم، به درستی تشخیص درد نوروپاتیک.
با این حال، برخی از ابزارهای تشخیصی وجود دارد که ممکن است برای ارزیابی درد نوروپاتی به پزشکان کمک کند. برای شروع، مطالعات هدایت عصب و پتانسیل های حسی شناختی می توانند میزان آسیب به مسیرهای حسی، اما نه غم انگیز، را از طریق نظارت بر پاسخ های نوروفیزیولوژیک به محرک های الکتریکی شناسایی و اندازه گیری کنند. علاوه بر این، تست های حسی سنجی کمی در واکنش به محرک های خارجی از شدت های مختلف با استفاده از تحریک به پوست می پردازند. حساسیت مکانیکی به محرک های لمسی با استفاده از ابزارهای تخصصی مانند موهای فون فریم، پینکر با سوزن های متصل، و همچنین حساسیت لرزش همراه با vibrameters و درد حرارتی با ترمود اندازه گیری می شود.
همچنین بسیار مهم است که ارزیابی جامع عصبی را برای شناسایی اختلالات حرکتی، حسی و اتوکی انجام دهیم. در نهایت، پرسشنامه های متعددی برای تشخیص درد نوروپاتیک در درد ناخوشی وجود دارد. بعضی از آنها فقط شامل پرسشنامه مصاحبه (مانند پرسشنامه نوروپاتیک و درد ID) می باشند، در حالی که دیگران شامل هر دو پرسش مصاحبه و آزمایش های فیزیکی (به عنوان مثال، ارزیابی نشانه های نوروپاتی و مقیاس علائم لیدز) و ابزار دقیق رمان، ارزیابی استاندارد درد، که شامل شش سوال مصاحبه و ده ارزیابی فیزیولوژیکی است.
روش های درمان درد نوروپاتیک
رژیم های دارویی به مکانیسم درد نوروپاتی کمک می کنند. با این حال، هر دو درمان فارماکولوژیک و غیر فارماکولوژیک تقریبا نیمی از بیماران، تسکین کامل یا جزئی را ارائه می دهند. بسیاری از توصیفات مبتنی بر شواهد نشان می دهد که استفاده از مخلوط دارو و یا دارو برای کارکردن به حداکثر مکانیزم ممکن است. اکثریت مطالعات بیشتر به بررسی نورالژی پس از هرپس و نوروپاتی های دردناک دیابتی محسوب می شوند، اما نتایج ممکن است در همه موارد درد نوروپاتیک نباشد.
داروهای ضد افسردگی
داروهای ضد افسردگی سطوح سیناپسی سروتونین و نوراپی نفرین را افزایش می دهند و درنتیجه اثر سیستم متابولیسم نزولی همراه با درد نوروپاتیک را افزایش می دهد. آنها اصلی ترین درمان درد نوروپاتیک بوده اند. اقدامات ضددردی ممکن است مربوط به انسداد مجدد نورآدرنالین و دوپامین باشد که احتمالا افزایش مهار نزولی، انطباق NMDA-گیرنده و انسداد کانال سدیم را افزایش می دهد. ضد افسردگیهای سه حلقه ای مانند TCAs؛ به عنوان مثال، آمیتریپتیلین، ایمیپرامین، نوریتیپتیلین و دوکسپین، در برابر درد مداوم درد یا سوزش همراه با درد خود به خودی قوی هستند.
داروهای ضد افسردگی سه حلقه ای به طور قابل توجهی برای درد نوروپاتیک نسبت به بازدارنده های بازجذب سروتونین خاص یا SSRI ها مانند فلوکستین، پاروکستین، سرترالین و سیتالوپرام اثبات شده است. ممکن است دلیل این باشد که جذب سرتوتونین و نور اپی نفرین را مهار می کنند، در حالی که SSRI ها فقط جذب سرتوتون را مهار می کنند. داروهای ضد افسردگی سه حلقه ای می توانند عوارض جانبی ناخوشایند، از جمله تهوع، سردرگمی، بلوک های هدایت قلبی، تاکی کاردی و آریتمی های بطنی داشته باشند. آنها همچنین می توانند سبب افزایش وزن، کاهش آستانه تشنج و فشار خون ارثا استاتیک شوند. سه حلقه چپ باید با مراقبت در سالمندان استفاده شود، که به خصوص عوارض جانبی حاد آنها آسیب پذیر است. غلظت دارو در خون باید برای جلوگیری از مسمومیت در بیماران که متابولیزه کننده داروهای آهسته است، کنترل شود.
مهارکننده های بازجذب سروتونین و نوراپی نفرین، یا SNRI، یک کلاس جدید ضد افسردگی هستند. مانند TCA ها، به نظر می رسد که آنها برای درمان درد نوروپاتیک نسبت به SSRI ها موثرتر هستند زیرا آنها همچنین جذب مجدد نوراپینفرین و دوپامین را مهار می کنند. Venlafaxine در برابر پلیeneuropathies تسکین دهنده، مانند نوروپاتی دردناک دیابتی، به عنوان imipramine، در اشاره به TCA موثر است، و این دو به طور قابل توجهی بیشتر از پلاسبو است. به نظر می رسد SNRI ها مانند TCA ها فواید را مستقل از اثرات ضد افسردگی خود می دانند. عوارض جانبی شامل آرام بخش، سردرگمی، فشار خون بالا و سندرم قطع است.
داروهای ضد صرع
داروهای ضد صرع می تواند به عنوان درمان خط اول مخصوصا برای انواع خاصی از درد نوروپاتیک استفاده شود. آنها با تعدیل کانال های کلسیم و سدیم با ولتاژ، با بهبود اثرات مهاری GABA و مهار انتقال گلوتامین ارگانیک، عمل می کنند. داروهای ضد صرعی برای درد حاد مؤثر نیست. در موارد درد مزمن، داروهای ضد صرعی تنها در نورالژی تریمینال موثرند. کاربامازپین به طور معمول برای این وضعیت کار می کند. گاباپنتین، که با مهار عملکرد کانال کلسیم از طریق فعالیت های آگونیست در زیر واحد دلتا آلفا-2 کانال کلسیم عمل می کند، همچنین برای درد نوروپاتیک موثر است. با این حال، گاباپنتین به طور مرکزی عمل می کند و ممکن است باعث خستگی، سردرگمی و خواب آلودگی شود.
غیر اپیوئیدی ضد درد دار
کمبود داده های قوی با استفاده از داروهای ضد التهابی غیر استروئیدی یا NSAID ها در کاهش درد نوروپاتیک وجود دارد. این ممکن است به دلیل عدم وجود عنصر التهابی در کاهش درد باشد. اما آنها به عنوان جایگزین در درمان درد سرطانی به عنوان جایگزینی با مواد مخدر مورد استفاده قرار گرفته اند. هرچند، عوارض گزارش شده است، به ویژه در بیماران شدیدا ناتوان.
داروهای ضد افسردگی اپوئیدی
مسکنهای ضد افسردگی در زمینه کاهش درد نوروپاتی موضوع بحث زیادی هستند. آنها با مهار امواج درد مرکزی صعودی عمل می کنند. به طور سنتی، درد نوروپاتیک قبلا مشاهده شده است به مقاومت در برابر opioid، که در آن opioids روش های مناسب تر برای انواع درد ناخوشایندی کرونری و جسمی است. بسیاری از پزشکان به دلیل نگرانی در مورد سوء مصرف مواد، اعتیاد و مسائل قانونی، از مصرف مواد مخدر برای درمان درد نوروپاتی استفاده می کنند. اما، محاکمه های زیادی وجود دارد که داروهای ضد افسردگی را برای موفقیت به دست آورده اند. اگزوكودون برای تسکین درد، آلدوینیا، بهبود خواب و معلولیت بهتر از پلاسبو بود. مواد مخدر کنترل شده آزاد، با توجه به برنامه ریزی شده، برای بیماران با درد ثابت برای تشویق سطح ثابت بیهوشی، جلوگیری از نوسانات گلوکز خون و جلوگیری از عوارض جانبی همراه با دوز بالاتر توصیه می شود. به طور گسترده ای، آماده سازی دهان به دلیل سهولت استفاده بیشتر و هزینه مقرون به صرفه می باشد. داروهای ترشحی پوستی، تزریقی و رکتال به طور کلی در بیمارانی که نمی توانند داروهای خوراکی را تحمل کنند استفاده می شود.
بیحسی موضعی
اعتیاد به مواد مخدر در نزدیکی جذاب است زیرا به لطف اقدام منطقه ای آنها عوارض جانبی کمتری دارند. آنها با تثبیت کانال های سدیم در آکسون های نورون های درجه اول محیطی عمل می کنند. آنها بهتر کار می کنند اگر فقط آسیب عصب جزئی و کانال های سدیم اضافی جمع آوری شده است. موضعی لیدوکائین بهترین نماینده دوره ای برای درد نوروپاتیک است. به طور خاص، استفاده از این پلات لیدوکائین 5 درصد برای نورالژی پس از هرپس، باعث تایید آن توسط FDA شده است. این پچ به نظر می رسد بهترین کار را در زمانی که آسیب دیده است، اما حفظ عملکرد سیستم عصبی محیطی از تخریب درماتومی که به عنوان آلدینیا نشان داده می شود. این باید مستقیما روی ناحیه علامت گذاری شده برای ساعت 12 تنظیم شود و برای یک ساعت دیگر 12 حذف شود و ممکن است برای سال ها به این روش استفاده شود. علاوه بر واکنش های موضعی پوست، اغلب به خوبی توسط بسیاری از بیماران مبتلا به درد نوروپاتی تحمل می شود.
داروهای متفرقه
کلونیدین، آگونیست آلفا-2، در تعدادی از بیماران مبتلا به نوروپاتی محیطی دیابتی موثر بوده است. کانابینوئید ها نقش مهمی در مدولاسیون درد در مدل های حیوانی ایفا می کنند و شواهدی از اثربخشی آن را تجمع می دهد. آگونیست های انتخابی CB2 سرکوب کننده هیپرالژیزیا و آلدوینیا هستند و آستانه های ناخوشایند را بدون ایجاد التیام بخش نرمال می کنند.
مدیریت درد مداخله
درمان های تهاجمی ممکن است برای بیماران مبتلا به درد نوروپاتیک قابل تحمل باشد. این درمان ها شامل تزریق اپیدورال یا پروینرژیک بیحسکننده های موضعی یا کورتیکواستروئیدها، تسریع در آوردن داروهای اپیدورال و اینتراکتال و افزودن محرک های نخاعی می باشد. این رویکردها برای بیماران مبتلا به درد نوروپاتیک مزمن قابل پیشگیری است که از نظر مدیریت پزشکی محافظه کار شکست خورده اند و همچنین ارزیابی روان شناختی کامل را تجربه کرده اند. در مطالعه کیم و همکاران، نشان داده شد که محرک نخاعی در درمان درد نوروپاتیک ریشه عصب موثر است.
دکتر الکس جیمنز Insight
با درد نوروپاتیک علائم درد مزمن به علت آسیب رساندن، غیرفعال بودن یا آسیب رسیدن به الیاف عصبی رخ می دهند، که معمولا با آسیب یا آسیب بافتی همراه است. در نتیجه، این فیبرهای عصبی می توانند سیگنال درد نادرست را به قسمت های دیگر بدن ارسال کنند. اثرات درد نوروپاتیک ناشی از آسیب های نخاعی عصبی شامل تغییر در عملکرد عصب در محل آسیب و در مناطق اطراف آسیب است. شناخت پاتوفیزیولوژی درد نوروپاتیک برای بسیاری از متخصصان مراقبت های بهداشتی هدف بوده است تا به طور موثری بهترین روش درمان را برای کمک به مدیریت و بهبود علائم آن ها تعیین کند. از استفاده از داروها و یا داروها، مراقبت از مراقبت از کایروپراکتیک، ورزش، فعالیت بدنی و تغذیه، ممکن است از روشهای مختلف درمان برای کمک به کاهش درد نوروپاتیک برای نیازهای هر فرد استفاده شود.
مداخلات اضافی برای درد نوروپاتیک
بسیاری از بیماران مبتلا به درد نوروپاتیک به دنبال گزینه های مکمل و جایگزین برای درمان درد نوروپاتیک هستند. دیگر رژیم های شناخته شده برای درمان درد نوروپاتیک شامل طب سوزنی، تحریک الکتریکی عصبی از طریق پوست، تحریک الکتریکی عصبی پوستی، درمان رفتاری شناختی، تصاویر حرکتی درجه بندی شده و درمان حمایتی و ورزش است. با این وجود، مراقبت از کایروپراکتیک یک رویکرد شناخته شده درمان جایگزین است که معمولا برای کمک به درمان درد نوروپاتی استفاده می شود. مراقبت های کایروپراکتیک همراه با فیزیوتراپی، ورزش، تغذیه و تغییرات شیوه زندگی می تواند در نهایت باعث کاهش علائم درد نوروپاتیک شود.
مراقبت کایروپراکتیک
شناخته شده است که یک برنامه مدیریت جامع برای مبارزه با اثرات درد نوروپاتیک بسیار مهم است. به این ترتیب، مراقبت از مراقبت از کایروپراکتیک یک برنامه درمان جامع است که می تواند در پیشگیری از مشکلات بهداشتی مرتبط با آسیب عصبی موثر باشد. مراقبت های کایروپراکتیک به بیماران با بسیاری از بیماری های مختلف کمک می کند، از جمله کسانی که درد نوروپاتیک دارند. بیماران مبتلا به درد نوروپاتیک اغلب از داروهای غیر استروئیدی ضد التهابی یا داروهای ضد التهابی مانند Ibuprofen یا داروهای ضد درد استفاده می کنند تا به کاهش درد نوروپاتیک کمک کنند. این ممکن است ثابت موقت را فراهم کند اما برای کنترل درد نیاز به استفاده مداوم دارد. این به طور مداوم به عوارض جانبی مضر و در شرایط شدید، وابستگی به داروهای تجویزی کمک می کند.
مراقبت از کایروپراکتیک می تواند به بهبود علائم درد نوروپاتیک کمک کند و ثبات را بدون این نواقص افزایش دهد. یک رویکرد مانند مراقبت از مراقبت از دهان و دندان، یک برنامه فردی طراحی شده برای مشخص کردن علت اصلی این موضوع است. از طریق استفاده از تنظیمات ستون فقرات و دستکاری دستی، یک جراح زیبایی می تواند به دقت هر اختلال ستون فقرات یا زیر شکمی که در طول طول ستون فقرات یافت می شود، که می تواند عواقب ناخوشایند عصب را از طریق اصلاح ستون فقرات کاهش دهد. بازسازی یکپارچگی ستون فقرات برای حفظ عملکرد سیستم عصبی مرکزی ضروری است.
یک فست فود کننده همچنین می تواند درمان طولانی مدت در جهت بهبود رفاه عمومی شما باشد. علاوه بر تنظیمات ستون فقرات و دستکاری دستی، یک پزشک ممکن است توصیه های تغذیه ای مانند تجویز یک رژیم غذایی حاوی آنتی اکسیدان ها را ارائه دهد یا آنها بتوانند یک برنامه فیزیوتراپی یا برنامه ورزشی برای مبارزه با اضطراب درد عصب طراحی کنند. یک وضعیت درازمدت مستلزم درمان طولانی مدت است و در این ظرفیت یک متخصص مراقبت های بهداشتی که متخصص جراحات و / یا شرایطی است که بر سیستم عضلانی اسکلتی و عصبی مانند پزشک متخصص کایروپراکتیک یا هیپراکتر متخصص است، ممکن است ارزشمند باشد زیرا آنها کار می کنند برای اندازه گیری تغییرات مطلوب در طول زمان.
روشهای درمانی فیزیکی، ورزش و نمایش جنبشی برای درمان درد نوروپاتیک مفید است. مراقبت های کایروپراکتیک همچنین سایر روش های درمان را ارائه می دهد که ممکن است برای مدیریت یا بهبود درد نوروپاتی مفید باشد. به عنوان مثال، لیزر سطح پایین یا LLLT، به عنوان یک درمان برای درد نوروپاتیک اهمیت فوق العاده ای به دست آورده است. با توجه به مطالعات مختلف تحقیقاتی، نتیجه گرفتند که LLLT تاثير مثبت بر کنترل درد زخم بر درد نوروپاتيک دارد، اما برای تعريف پروتکل های درماني که در آنها اثرات ليزر درماني پايين در درمان درد نوروپاتيک خلاصه مي شود، مطالعات بيشتري در اين زمينه انجام شده است.
مراقبت های کایروپراکتیک همچنین شامل توصیه های تغذیه ای است که می تواند به کنترل علائم مرتبط با نوروپاتی دیابتی کمک کند. در طی یک مطالعه تحقیقاتی، یک رژیم غذایی مبتنی بر گیاه کم چرب به منظور بهبود کنترل گلیسمی در بیماران مبتلا به دیابت نوع 2 نشان داده شد. بعد از حدود هفته های آزمایش 20، افرادی که در معرض تغییرات در وزن بدن خود قرار گرفته اند و میزان هدایت الکتریکی پوست در پا بهبود یافته است، با مداخله بهبود یافته است. تحقیقات انجام شده نشان می دهد که مقدار بالقوه در مداخلات رژیم غذایی کم چربی برای مبتلایان به نوروپاتی دیابتی پیشنهاد شده است. علاوه بر این، مطالعات بالینی نشان می دهد که کاربرد خوراکی منیزیم L-threonate قادر به جلوگیری و همچنین بهبود نقص حافظه همراه با درد نوروپاتیک است.
مراقبت های کایروپراکتیک همچنین می تواند راهکارهای درمان اضافی را برای ارتقاء بازسازی عصب ارائه دهد. به طور مثال، بهبود بازسازی آکسون پیشنهاد شده است تا به بهبود بهبود عملکردی پس از آسیب عصبی محیطی کمک شود. با توجه به مطالعات انجام شده اخیر، تحریک الکتریکی، همراه با فعالیت های ورزشی و یا فعالیت های فیزیکی، باعث بهبود بازسازی عصب پس از تأخیر در تعمیر عصب در انسان و موش می شود. هر دو محرک الکتریکی و ورزش در نهایت مصمم به درمان تجربی برای آسیب عصبی محیطی بودند که ظاهرا آماده انتقال به استفاده بالینی بودند. برای تعیین کامل اثرات این در بیماران مبتلا به درد نوروپاتیک، ممکن است مطالعات بیشتری انجام شود.
نتیجه
درد نوروپاتیک یک نهاد چندوجهی است که هیچ دستورالعمل خاصی برای مراقبت از آن وجود ندارد. بهتر است با استفاده از یک رویکرد چند رشته ای مدیریت شود. مدیریت درد مستلزم ارزیابی مداوم، آموزش بیمار، اطمینان از پیگیری بیمار و اطمینان خاطر است. درد نوروپاتیک یک بیماری مزمن است که گزینه بهترین درمان را چالش برانگیز می کند. درمان فردی شامل در نظر گرفتن تأثیر درد بر بهزیستی، افسردگی و ناتوانی های فرد همراه با آموزش و ارزیابی مداوم است. مطالعات درد نوروپاتیک، هم در سطح مولکولی و هم در مدل های حیوانی، نسبتاً جدید اما بسیار امیدوارکننده است. پیشرفتهای زیادی در زمینههای اساسی و بالینی درد نوروپاتیک پیشبینی میشود، از این رو راههایی برای روشهای درمانی بهبودیافته یا جدید برای این وضعیت ناتوانکننده باز میشود. دامنه اطلاعات ما محدود به کایروپراکتیک و همچنین آسیب ها و شرایط ستون فقرات است. برای بحث در مورد موضوع، لطفاً از دکتر خیمنز بپرسید یا با ما تماس بگیرید915-850-0900.
دکتر الکس جیمنز سرپرستی می کند
موارد اضافی: درد پشت
درد پشت یکی از شایع ترین علل ناتوانی و از دست رفته روز در کار در سراسر جهان است. به عنوان حقيقت، درد پشتي به عنوان دومين شايعترين دلايل بازديد پزشکي، نسبت به عفونت فوقاني تنفسي کمتر است. تقریبا 80 درصد از جمعیت حداقل یک بار در طول زندگی خود نوعی کمر درد را تجربه خواهند کرد. ستون فقرات یک ساختار پیچیده ای است که از استخوان ها، مفاصل، لیگامان ها و ماهیچه ها در میان دیگر بافت های نرم تشکیل شده است. به همین دلیل، آسیب و / یا شرایط شدید، مانند دیسک های فتق دیسک، در نهایت می تواند منجر به علائم درد پشت شود. آسیب های ورزشی یا آسیب های ناشی از تصادفات خودرو اغلب اغلب علت درد پشت هستند، اما گاهی اوقات ساده ترین حرکات می توانند نتایج دردناکی داشته باشند. خوشبختانه، گزینه های درمان جایگزین، مانند مراقبت از کیهان پراکسی، می تواند به کاهش درد در استفاده از تنظیمات ستون فقرات و دستکاری دست کمک کند و در نهایت بهبود تسکین درد را کاهش دهد.
افسردگی اضطراب دردveهمه افراد درد را تجربه کرده اند ، با این حال ، کسانی مبتلا به افسردگی ، اضطراب یا هر دو هستند. این را با درد ترکیب کنید و درمان آن بسیار شدید و دشوار می شود. افرادی که از افسردگی ، اضطراب یا هر دو رنج می برند بیشتر از سایر افراد دچار درد شدید و طولانی مدت می شوند.
راه اضطرابافسردگی و درد همپوشانی یکدیگر را در بیماران مزمن و برخی از سندرم های درد ناشنوا مشاهده می کنند، یعنی کمردرد، سردرد، درد عصبی و فیبرومیالژیا. اختلالات روانپزشکی به شدت درد و همچنین افزایش خطر ابتلا به ناتوانی کمک می کند.
افسردگی: A (اختلال افسردگی اساسی یا افسردگی بالینی) یک اختلال خلقی شایع اما جدی است. این علائم شدیدی ایجاد می کند که بر احساس ، تفکر و نحوه کنترل فعالیتهای روزمره ، یعنی خواب ، غذا و کار تأثیر می گذارد. برای تشخیص افسردگی ، علائم باید حداقل به مدت دو هفته وجود داشته باشد.
حال و هوای مداوم غمگین ، مضطرب یا خسته کننده.
احساس ناامیدی، بدبینانه.
تحریک پذیری.
احساس گناه، بی ارزش بودن یا بی کفایتی.
از دست دادن علاقه یا لذت در فعالیت ها.
کاهش انرژی یا خستگی.
حرکت یا صحبت کردن به آرامی.
احساس بی قراری و مشکل در نشستن آرام.
دشواری تمرکز، یادآوری یا تصمیم گیری.
مشکل خواب ، بیدار شدن در صبح زود و خوابیدن بیش از حد.
تغییرات اشتها و وزن
افکار مرگ یا خودکشی و یا اقدام به خودکشی.
درد یا درد، سردرد، گرفتگی، و یا مشکلات گوارشی بدون علت جسمی روشن و یا که با درمان آسان نیست.
همه افراد افسرده همه علائم را تجربه نمی کنند. برخی فقط چند علامت را تجربه می کنند در حالی که برخی دیگر ممکن است چندین علامت را تجربه کنند. چندین علائم پایدار علاوه بر روحیه پایین نیز وجود داردضروری برای تشخیص افسردگی اساسی. شدت و دفعات علائم همراه با مدت زمان بستگی به فرد و بیماری خاص آنها دارد. علائم نیز بسته به مرحله بیماری ممکن است متفاوت باشد.
افسردگی اضطراب درد
اهداف:
رابطه چیست؟
نوروفیزیولوژی در پشت آن چیست؟
پیامدهای مرکزی چیست؟
تغییرات مغزی در درد
شکل 1 مسیرهای مغز، مناطق و شبکه های درگیر در درد حاد و مزمن
دیویس، KD و همکاران. (2017) آزمایش های تصویربرداری مغز برای درد مزمن: مسائل پزشکی و قانونی و اخلاقی و توصیه های Nat. نوئور. doi: 10.1038 / nrneurol.2017.122
درد، اضطراب و افسردگی
نتیجه:
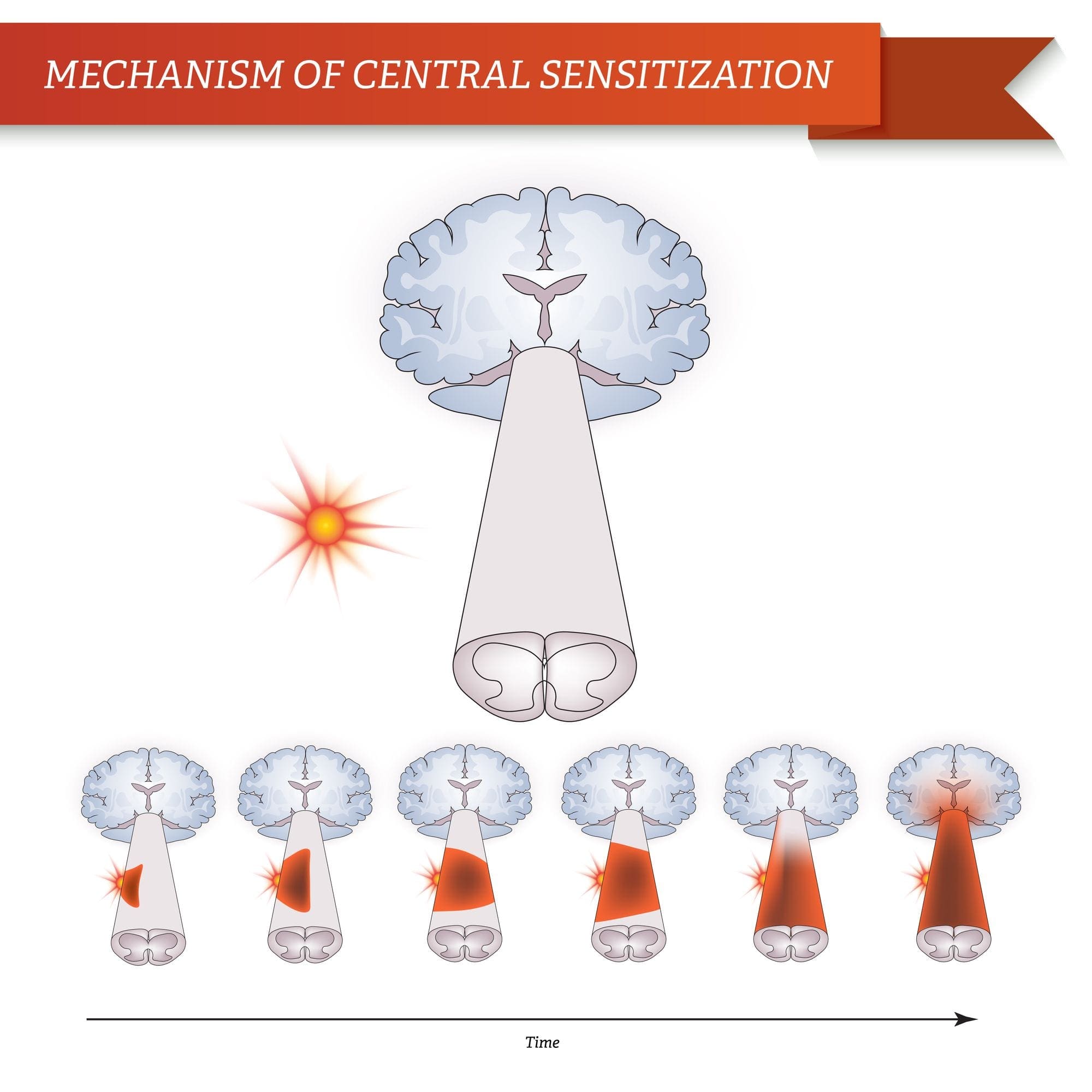
درد، به خصوص مزمن با افسردگی و اضطراب همراه است
مکانیسم های فیزیولوژیکی که منجر به اضطراب و افسردگی می شوند، می توانند به صورت ماهیتا چندگانه باشند
هنگامی که سیستم حسی تحت تأثیر آسیب یا بیماری قرار می گیرد، اعصاب درون آن سیستم نمی توانند به خوبی برای انتقال احساسات و احساسات به مغز کار کنند. این اغلب به احساس بیحسی یا عدم حس اعتماد به نفس کمک می کند. با این حال، در موارد خاص، زمانی که این سیستم آسیب دیده، مردم ممکن است درد در ناحیه آسیب دیده را تجربه کنند.
درد به طور ناگهانی شروع نمی شود و یا به سرعت حل می شود این یک درد مزمن شرایطی که منجر به علائم درد مداوم می شود. برای اکثر افراد، شدت علائم آنها ممکن است در طول روز از بین برود و از بین برود. گرچه درد نوروپاتي در ارتباط با مسائل بهداشتي عصب محيطي مانند نوروپاتي ناشي از ديابت يا تنگي ستون فقرات، جراحات مغز و نخاع مي تواند منجر به درد مزمن نوروپاتي شود. درد نوروپاتیک نیز به عنوان درد عصب شناخته شده است.
درد نوروپاتیک ممکن است در مقایسه با درد ناخوشایند باشد. درد نوروپاتیک به هیچ یک از شرایط خاص یا محرک خارج نمی رسد، بلکه این علائم فقط به این دلیل رخ می دهند که سیستم عصبی ممکن است بر آن کار نکند. در واقع، افراد همچنین می توانند درد نوروپاتیک را تجربه کنند، حتی زمانی که قسمت اعصاب آسیب دیده یا آسیب دیده واقع نمی شود. این وضعیت درد فانتوم درد است که ممکن است در افراد بعد از قطع عضو رخ دهد.
درد ناخوشایندی معمولا حاد است و در پاسخ به یک شرایط خاص مانند زمانیکه کسی آسیب ناگهانی را تجربه می کند، مثل شلاق زدن انگشت با یک چکش یا پا کردن پا در حین راه رفتن پابرهنه. علاوه بر این، درد ناخوشی پس از درمان سایت بهبود می یابد. این بدن حاوی سلول های عصبی تخصصی است که به نام های نوکسیپتور شناخته می شود که باعث تشخیص محرک های خطرناکی می شود که می توانند به بدن آسیب برسانند، مانند گرما شدید یا سرد، فشار، تکان دادن و قرار گرفتن در معرض مواد شیمیایی. سپس این سیگنال های هشدار دهنده در امتداد سیستم عصبی به مغز منتقل می شود و موجب درد ناخوشی می شود.
عوامل خطر برای درد نوروپاتی چیست؟
هر چیزی که باعث کمبود عملکرد در سیستم عصبی حسی می شود می تواند به درد نوروپاتیک منجر شود. به همین ترتیب، مسائل بهداشتی عصب از سندرم تونل کارپ، و یا شرایط مشابه، در نهایت باعث درد نوروپاتیک می شود. تروما، منجر به آسیب عصبی، ممکن است منجر به درد نوروپاتیک شود. سایر شرایطی که می توانند افراد را در ایجاد درد نوروپاتیک مستعد کنند عبارتند از: دیابت، کمبود ویتامین، سرطان، اچ آی وی، سکته مغزی، مولتیپل اسکلروز، زونا و حتی برخی از درمان های سرطان.
علل درد نوروپاتیک چیست؟
علل زیادی وجود دارد که افراد ممکن است درد نوروپاتیک ایجاد کنند. اما در یک سطح سلولی، یک توضیح، افزایش گسیل گیرنده های خاصی است که نشان دهنده درد، و همچنین توانایی کاهش توانایی اعصاب برای تعدیل این سیگنال ها است، منجر به احساس درد ناشی از منطقه آسیب دیده می شود. علاوه بر این، در ناحیه نخاعی، منطقه ای که علائم دردناک را تحریک می کند، با تغییرات متناوب در هورمون ها و از دست دادن اجسام متحرک متحرک بازسازی می شود. این تغییرات منجر به درک درد در غیاب تحریک خارجی می شود. در مغز، توانایی بلوکه شدن درد ممکن است پس از آسیب، مانند سکته مغزی یا آسیب از آسیب، تحت تأثیر قرار گیرد. با گذشت زمان، آسیب سلولی اضافی اتفاق می افتد و احساس درد ادامه دارد. درد نوروپاتیک نیز به دیابت، مصرف الکل مزمن، سرطانهای خاص، کمبود ویتامین B، بیماری، سایر بیماریهای مرتبط با عصب، سموم و داروهای خاص مرتبط است.
علائم درد نوروپاتیک چیست؟
بر خلاف سایر شرایط عصبی، شناسایی درد نوروپاتیک می تواند چالش برانگیز باشد. با این حال، ممکن است چندین سیگنال عینی وجود داشته باشند. متخصصان بهداشت و درمان باید رمزگشایی و ترجمه مجموعه ای از کلمات که بیماران استفاده می شود برای توصیف درد خود را. بیماران ممکن است علائم خود را به عنوان تیز، خفیف، گرم، سرد، حساس، خارش، عمیق، خارش، سوزش، در میان انواع مختلف توصیف شرایط توصیف کنند. علاوه بر این، برخی از بیماران ممکن است درد را از طریق لمس یا فشار نور تجربه کنند.
در تلاش برای شناسایی میزان درد بیماران در معرض خطر، اغلب مقادیر مختلف استفاده می شود. از بیماران خواسته می شود درد خود را با توجه به یک مقیاس بصری یا نمودار عددی رتبه بندی کنند. بسیاری از نمونه های مقیاس درد وجود دارد، مانند مثال زیر نشان داده شده است. اغلب تصاویری از چهره هایی که انواع سطوح مختلف درد را نشان می دهند ممکن است مفید باشد وقتی که افراد زمان دشوار توصیف مقدار درد که در حال تجربه هستند.
درد مزمن و سلامت روان
برای بسیاری از افراد، درد مزمن ممکن است به درد محدود نشود. همچنین ممکن است بر وضعیت ذهنی آنها تأثیر منفی داشته باشد. مطالعات جدید انجام شده توسط دانشمندان در دانشگاه شمال غرب در شیکاگو می تواند توضیح دهد که چرا افراد مبتلا به درد مزمن نیز با مشکلات بهداشتی ظاهرا غیر مرتبط، مانند افسردگی، استرس، عدم خواب و تمرکز دشواری رنج می برند.
ارزیابی نشان داد که افراد مبتلا به درد مزمن، مناطق مختلف مغز را نشان می دهند که همیشه فعال هستند، به طور خاص، منطقه ای که با خلق و ذهن توجه می شود. این اقدام مداوم ارتباطات عصبی را از مغز باز می کند و بیماران مزمن درد را در معرض خطر بیشتری برای مشکلات روحی قرار می دهد. محققان پیشنهاد کرده اند که دریافت سیگنال های درد به طور دائم می تواند موجب عقب ماندگی ذهنی شود که بر ذهن تاثیر می گذارد. احیای مجدد مغز آنها را مجبور می کند تا منابع ذهنی متنوعی را برای مقابله با کارهای روزمره، از ریاضیات، به یاد آوردن یک لیست خرید، خوشحال کنند.
اتصال درد و مغز به خوبی ثبت شده است، حداقل عجیب و غریب، و بسیاری از متخصصان مراقبت های بهداشتی می گویند که آنها از دست اول به عنوان حالت روانی بیمار می تواند در هنگام سقوط درد مزمن تحمل کند. اشتباهات در مورد ارتباط درد و مغز ممکن است ناشی از فقدان شواهدی باشد مبنی بر اینکه درد در اندازه گیری و تأثیر پایدار بر مغز دارد. محققان انتظار دارند که با تحقیقات بیشتری در مورد مکانیسم هایی که چگونه درد مزمن افراد را بیشتر در معرض اختلالات خلقی قرار می دهد، مردم می توانند توانایی مدیریت بهتر رفاه عمومی خود را داشته باشند.
فرهنگ و درد مزمن
با این حال، بسیاری از چیزها به شیوۀ تجربه و بیان درد کمک می کنند، اما اخیرا توسط محققان پیشنهاد شده است که فرهنگ به طور مستقیم به بیان درد مرتبط است. ارزش های تربیتی و اجتماعی ما بر چگونگی بیان درد و همچنین طبیعت، شدت و طول خود ما تاثیر می گذارد. با این حال، این متغیرها به اندازه ارزشهای اجتماعی-روانشناختی مانند سن و جنس واضح نیستند.
تحقیقات نشان می دهد که درد مزمن یک فرایند چند وجهی است و همپوشانی همزمان بین پاتوفیزیولوژی، عوامل شناختی، عاطفی، رفتاری و جامعه شناختی به آنچه که تجربه درد مزمن نامیده می شود، خلاصه می شود. به نظر می رسد که درد مزمن در بیماران از فرهنگ ها و قومیت های مختلف متفاوت است.
برخی از فرهنگ ها بیان درد را تشویق می کنند، به ویژه در مدیترانه جنوبی و خاورمیانه. افراد دیگر آن را سرکوب می کنند، همانطور که در بسیاری از درس های ما بچه ها در مورد رفتار شجاعانه و نه گریه. درد به عنوان بخشی از تجربه انسان شناخته شده است. ما معتقدیم که ارتباط در مورد درد به طور یکپارچه مرزهای فرهنگی متقابل است. اما افراد مبتلا به درد به شیوهای رفتار می کنند که تمدنشان آموزش می دهند تا تجربه و ابراز درد کنند.
هر دوی این افراد در درد و متخصصین مراقبت های بهداشتی مشکالت ارتباط درد در بین مرزهای قومی را تجربه می کنند. در یک موضوع مانند درد، که در آن ارتباطات موثر می تواند پیامد های گسترده ای برای مراقبت های پزشکی، کیفیت زندگی و بقاء بالقوه داشته باشد، نقش فرهنگ در ارتباط با درد هنوز تحت بررسی نیست. درد مداوم یک رویکرد چند بعدی و یکپارچه است که با درهم آمیختن و همبستگی با عوامل بیولوژیک و روان شناختی شکل می گیرد. دانستن نقطه اوج این عوامل برای درک تفاوت های ظاهری و مدیریت آن ضروری است.
چگونه درد نوروپاتیک تشخیص داده می شود؟
تشخیص درد نوروپاتیک مبتنی بر ارزیابی اضافی از تاریخ فرد است. اگر مشکوک به آسیب عصبی پایه باشد، تجزیه و تحلیل اعصاب و آزمایشات ممکن است توجیه شود. شایعترین وسیله برای ارزیابی اینکه آیا یک عصب مجروح شده یا نه، استفاده از دارو الکترودیاژنتیک است. این فوق تخصص پزشکی با استفاده از تکنیک های مطالعات هدایت عصبی با الکترومیلیوگرافی (NCS / EMG) است. ارزیابی بالینی ممکن است شواهدی از از دست دادن کار را نشان دهد و می تواند شامل ارزیابی لمس نور، قابلیت تمایز تیز از درد مبهم و توانایی تشخیص دما و همچنین ارزیابی ارتعاش باشد.
پس از انجام یک معاینه بالینی کامل، تجزیه و تحلیل الکترودهای تشخیصی می تواند برنامه ریزی شود. این مطالعات توسط متخصص مغز و اعصاب انجام می شود. اگر نوروپاتی مشکوک باشد، شکار برای علل برگشت پذیر باید انجام شود. این می تواند شامل عملکرد خون برای کمبود ویتامین یا مشکلات تیروئید و مطالعات تصویربرداری برای حذف یک ضایعه ساختاری که بر نخاع تاثیر می گذارد. بسته به نتایج این آزمایش، ممکن است وسیله ای برای کاهش شدت نوروپاتی و احتمالا کاهش درد که بیمار در معرض آن قرار گیرد وجود داشته باشد.
متأسفانه، در بسیاری از موارد حتی کنترل خوبی از علت اصلی نوروپاتی، درد نوروپاتیک را معکوس نمیکند. این معمولا در بیماران مبتلا به نوروپاتی دیابتی دیده می شود. در موارد نادر، ممکن است نشانه هایی از تغییرات در الگوی رشد پوست و مو در منطقه آسیب دیده وجود داشته باشد. این تغییرات ممکن است با تغییرات جگر همراه باشد. در صورت وجود، این تغییرات می تواند به تعیین احتمال وجود درد نوروپاتیک مرتبط با یک بیماری که به عنوان سندرم درد پیچیده منطقه ای شناخته می شود، کمک کند.
دکتر الکس جیمنز Insight
درد نوروپاتیک یک بیماری مزمن درد است که عموما با آسیب مستقیم یا آسیب به سیستم عصبی یا اعصاب مرتبط است. این نوع درد از درد ناخوشی یا احساس درد معمولی متفاوت است. درد ناخوشی یک احساس حاد یا ناگهانی درد است که باعث می شود سیستم عصبی بلافاصله بعد از تروما سیگنال های درد را ارسال کند. اما با درد نوروپاتیک، بیماران ممکن است تیراندازی، سوزش درد بدون آسیب یا آسیب مستقیم داشته باشند. درک علل احتمالی درد نوروپاتیک بیمار در مقایسه با سایر انواع درد، می تواند به متخصصان مراقبت های بهداشتی کمک کند که روش های بهتر برای درمان درد مزمن را پیدا کنند.
درمان درد نوروپاتیک چیست؟
داروهای مختلف در تلاش برای درمان درد نوروپاتیک استفاده می شود. اکثریت این داروها از بین می روند، بدین معنی که دارو توسط FDA برای درمان بیماری های مختلف مورد تایید قرار گرفته و پس از آن به عنوان مفید برای درمان درد نوروپاتیک شناخته شده است. داروهای ضد افسردگی سه حلقه ای مانند آمیتریپتیلین، نوریتیپتیلین و دیزپارمین برای کنترل درد نوروپاتیک چندین ساله تجویز شده اند.
بعضی از افراد می بینند که این ممکن است در ارائه کمک به آنها بسیار موثر باشد. سایر داروهای ضد افسردگی نشان داده اند که برخی از امراض را ارائه می دهند. مهار کننده های بازجذب سروتونین انتخابی یا SSRI ها مانند پاروکستین و سیتالوپرام و سایر داروهای ضد افسردگی مانند ونلافاکسین و بوپروپیون در برخی از بیماران مورد استفاده قرار می گیرند. یکی دیگر از درمان های مکرر درد نوروپاتیک شامل داروهای ضد انعقاد، از جمله کاربامازپین، فنیتوین، گاباپنتین، لاموتریژین و دیگران است.
در موارد حاد نوروپاتی دردناک که به کارگروه خط اول پاسخ نمی دهند، داروهایی که معمولا برای درمان آریتمی های قلبی استفاده می شوند، ممکن است مفید باشد. با این حال، این می تواند به عوارض جانبی قابل توجهی منجر شود و اغلب باید از نزدیک تحت نظارت باشد. داروهایی که به طور مستقیم به پوست اعمال می شوند می توانند برای بعضی از بیماران منافع قابل توجهی داشته باشند. اشکال معمول استفاده شده عبارتند از لیدوکائین (در نوع پچ یا ژل) یا کپسایسین.
درمان درد نوروپاتیک وابسته به علت اصلی است. اگر علت برگشت پذیر باشد، اعصاب محیطی می تواند دوباره رشد کند و درد کاهش یابد؛ با این وجود، این کاهش درد ممکن است چند ماه تا چند سال طول بکشد. چندین گزینه دیگر درمان جایگزین، از جمله مراقبت از مراقبت از ماساژ درمانی و فیزیوتراپی، همچنین می تواند به منظور کمک به کاهش تنش و استرس در طول اعصاب، در نهایت کمک به بهبود علائم دردناک.
پیش آگهی درد نوروپاتیک چیست؟
بسیاری از افراد مبتلا به درد نوروپاتیک می توانند برخی از معیارهای کمک را دریافت کنند، حتی زمانی که درد آنها همچنان ادامه دارد. گرچه درد نوروپاتیک برای بیمار خطرناک نیست، حضور درد مزمن می تواند منفی بر کیفیت زندگی باشد. بیماران مبتلا به درد عصبی مزمن ممکن است از محرومیت از خواب یا اختلالات خلقی مانند افسردگی، اضطراب و استرس رنج ببرند، همانطور که قبلا در بالا ذکر شد. به علت آلوپسی ذاتی و عدم بازخورد حساسیتی، بیماران در معرض خطر ابتلا به آسیب یا عفونت قرار دارند و یا به طور ناخودآگاه باعث افزایش شدید آسیب کنونی می شوند. بنابراین، ضروری است که به مراقبت پزشکی فوری توجه کنید و از ایمنی و احتیاط اطمینان حاصل کنید.
آیا درد نوروپاتی می تواند مانع شود؟
بهترین راه برای جلوگیری از درد نوروپاتی، جلوگیری از پیشرفت یا پیشرفت نوروپاتی است. نظارت و تغییر گزینه های سبک زندگی، از جمله محدود کردن استفاده از الکل و تنباکو؛ حفظ وزن مناسب برای کاهش احتمال ابتلا به دیابت، بیماری های دژنراتیو مفصلی یا سکته؛ و داشتن فرم ارگونومیک عالی در محل کار یا هنگام انجام سرگرمی ها برای کاهش خطر آسیب استرس مکرر، استراتژی هایی برای کاهش احتمال ابتلا به نوروپاتی و درد بالقوه نوروپاتیک هستند. مطمئن شوید که در صورت بروز هر گونه علائم مرتبط با درد نوروپاتیک فوراً به دنبال مراقبت های پزشکی باشید تا با مناسب ترین رویکرد درمانی ادامه دهید. دامنه اطلاعات ما محدود به کایروپراکتیک و همچنین آسیب ها و شرایط ستون فقرات است. برای بحث در مورد موضوع، لطفاً از دکتر خیمنز بپرسید یا با ما تماس بگیرید915-850-0900.
دکتر الکس جیمنز سرپرستی می کند
موارد اضافی: درد پشت
درد پشت یکی از شایع ترین علل ناتوانی و از دست رفته روز در کار در سراسر جهان است. به عنوان حقيقت، درد پشتي به عنوان دومين شايعترين دلايل بازديد پزشکي، نسبت به عفونت فوقاني تنفسي کمتر است. تقریبا 80 درصد از جمعیت حداقل یک بار در طول زندگی خود نوعی کمر درد را تجربه خواهند کرد. ستون فقرات یک ساختار پیچیده ای است که از استخوان ها، مفاصل، لیگامان ها و ماهیچه ها در میان دیگر بافت های نرم تشکیل شده است. به همین دلیل، آسیب و / یا شرایط شدید، مانند دیسک های فتق دیسک، در نهایت می تواند منجر به علائم درد پشت شود. آسیب های ورزشی یا آسیب های ناشی از تصادفات خودرو اغلب اغلب علت درد پشت هستند، اما گاهی اوقات ساده ترین حرکات می توانند نتایج دردناکی داشته باشند. خوشبختانه، گزینه های درمان جایگزین، مانند مراقبت از کیهان پراکسی، می تواند به کاهش درد در استفاده از تنظیمات ستون فقرات و دستکاری دست کمک کند و در نهایت بهبود تسکین درد را کاهش دهد.
اگر سیستم حسی تحت تأثیر آسیب یا بیماری قرار گیرد ، اعصاب موجود در آن سیستم نمی توانند در انتقال احساس به مغز عمل کنند. این می تواند منجر به احساس بی حسی ، یا عدم احساس شود. در برخی موارد که سیستم حسی آسیب دیده است ، افراد می توانند در ناحیه آسیب دیده درد داشته باشند. درد نوروپاتیک به سرعت شروع نمی شود یا به سرعت پایان می یابد. این یک بیماری مزمن است که منجر به علائم آن می شود درد مداوم. برای بسیاری از افراد، شدت علائم می تواند در طول یک روز اتفاق بیفتد. تصور می شود که درد نوروپاتیک با مشکلات عصبی محیطی همراه است، به همین دلیل نوروپاتی ناشی از دیابت، تنگی نخاعی، آسیب مغز و نخاع می تواند منجر به درد مزمن نوروپاتیک شود.
درد عصبی
اهداف:
آن چیست؟
پاتوفیزیولوژی پشت آن چیست؟
علل چیست؟
برخی از مسیرها چیست؟
چگونه می توانیم آنرا حل کنیم؟
درد عصبی
درد ناشی از یک ضایعه اولیه یا اختلال در سیستم عصبی سموتوسنسسی است.
درد نوروپاتیک معمولا مزمن، دشوار است برای درمان و اغلب به درمان استاندارد ضد درد.
پاتوژنز درد نروپاتیک
مكانيزمهاي پرپهكار
پس از یک ضایعه عصبی محیطی، نورون ها حساس تر می شوند و تحریک پذیری غیر طبیعی و حساسیت به تحریک را افزایش می دهند
این به عنوان ... حساسیت محیطی شناخته شده است!
مکانیک مرکزی
به عنوان یک نتیجه از فعالیت در حال انجام خود به خود در ناحیه پیرامون، نورون ها فعالیت فعال پس زمینه را افزایش می دهند، زمینه های گیرنده بزرگ شده و پاسخ های بیشتر به امواج مضر، از جمله محرک های لمسی نرمال
این به معنای ...حساسیت مرکزی!
علل شایع
ضایعات یا بیماری های سیستم عصبی سموتوسنسوری می تواند منجر به انتقال تغییرات و اختلال در انتقال سیگنال های حسی به نخاع و مغز شود. شرایط مشترک با درد نوروپاتیک عبارتند از:
نورالژیستی پس از پیوند
نورالژی ترجیامین
رادیکولوپاتی دردناک
نوروپاتی دیابتی
عفونت HIV
جهش
قطع شدن
درد عصبی محیطی
سکته مغزی (به صورت درد پس از سکته مرکزی)
درد اندام فانتوم و واقعیت افزوده
درد فانتوم درد و AR
تزریق NEUROGENIC
اهداف:
آن چیست؟
پاتوفیزیولوژی پشت آن چیست؟
علل چیست؟
چگونه می توانیم آنرا حل کنیم؟
تزریق NEUROGENIC
التهاب نوروژنیک پاسخ التهابی موضعی ناشی از ناخوشایندی است که به واسوادالیته گفته می شود، افزایش نفوذ پذیری عروق، degranulation مشت سلولی و انتشار neuropeptides از جمله پپتید مرتبط با ژن SP و کلسی تونین (CGRP)
به نظر می رسد که نقش مهمی در پاتوژنز بیماری های متعدد از جمله میگرن، پسوریازیس، آسم، فیبرومیالژیا، اگزما، روده ای، دیستونی و حساسیت شیمیایی چندگانه ایفا می کند
علل شایع
مسیرهای متعددی وجود دارد که التهاب نوروژنیک ممکن است آغاز شود. این به خوبی مستند شده است، با استفاده از هر دو مدل حیوانات و نورون های جدا شده در آزمایشگاهی، کپسایسین، گرما، پروتون، برادیکینین و تريپتاز، رگولاتورهای جریان بالا از نفوذ کلسیم داخل سلولی هستند که منجر به انتشار نوروپپتیدی التهابی می شود. در مقابل تصور می شود که پروستاگلندین E2 و I2، سیتوکین ها، اینترلوکین-1، اینترلوکین-6 و فاکتور نکروز تومور باعث ایجاد انتشار عصبی خود نمی شوند، بلکه باعث تحریک نورون های حسی می شود و بنابراین آستانه برای شلیک را کاهش می دهد و موجب آزادی بیشتر نوروپپتیدها
در حالی که التهاب نوروژنیک به طور گسترده مورد مطالعه قرار گرفته است و به خوبی در بافت های محیطی مستند شده است، تا همین اواخر مفهوم التهاب نوروژنیک درون CNS عمدتا ناشناخته باقی مانده است. با توجه به ظرفیت التهاب نوروژنیک برای نفوذ پذیری عروق و منجر به ایجاد پدیده ادم، اکنون به طور گسترده ای برای پتانسیل آن برای نفوذپذیری BBB و ادم واژگونیک در مغز و نخاع تحت شرایط مختلف پاتولوژیک مورد بررسی قرار گرفته است.
همه ما به عنوان کودکان یاد گرفتیم که حواس های 5 وجود دارد: دید، طعم، صدا، بوی و لمس. چهار حس اول از ارگان های واضح و متمایز مانند چشم ها، جوانه های طعم، گوش ها و بینی استفاده می کنند، اما دقیقا چطور حس بدن را لمس می کنید؟ لمس روی تمام بدن، در داخل و خارج از بدن تجربه می شود. یک ارگان مجزا وجود ندارد که مسئول حس لمس است. در عوض، گیرنده های کوچک، یا انتهای عصبی، در سراسر بدن وجود دارد که حس لمس آن را می گیرد و سیگنال های مغز را با اطلاعات مربوط به نوع لمسی که رخ داده است، می فرستد. به عنوان جوانه طعم دهنده در زبان عطر و طعم تشخیص، mechanoreceptors غده در داخل پوست و سایر اندام ها است که احساس لمس را تشخیص. آنها به عنوان شناخته می شوند مکانیسم پذیرش زیرا آنها طراحی شده اند تا احساسات مکانیکی یا تفاوت های فشار را تشخیص دهند.
نقش مکانیسم گیرنده ها
فرد متوجه می شود که وقتی احساس مسئولیت برای کشف این حس خاص پیامی را به مغز ارسال می کند احساس حس می کنند که یک عضو اصلی است که تمام اطلاعات را پردازش می کند و مرتب می کند. پیام ها از طریق تمام سیم های مغز به مغز ارسال می شوند. هزاران نورون کوچک وجود دارد که به تمام نقاط بدن انسان متصل هستند و در انتهای بسیاری از این نورون ها مکانیس گیر هستند. برای نشان دادن آنچه اتفاق می افتد هنگامی که یک جسم را لمس می کنید، از مثال استفاده خواهیم کرد.
پیش بینی یک زمین پشه در بازوی شما. فشار این حشره، به طوری که نور، مکانیس گیرنده ها را در آن منطقه خاص بازوی تحریک می کند. این مکانیس گیرها پیامی را در امتداد نورون که به آن وصل شده اند ارسال می کنند. نورون تمام راه را به مغز متصل می کند، که پیامی را دریافت می کند که چیزی در بدن شما در محل دقیق مکانیسم گیرنده خاصی که پیام را ارسال می کند، لمس می شود. مغز با این توصیه عمل می کند. شاید به چشمان خود نگاه کند تا ناحیه بازوی که امضای را تشخیص دهد نگاه کند. و هنگامی که چشم ها به مغز می گویند که یک پشه بر روی بازوی وجود دارد، مغز ممکن است دست به سرعت آن را بچرخاند. این کار چگونه کار می کند. هدف مقاله زیر نشان دادن و همچنین به طور مفصل در مورد سازماندهی عملکردی و مولکولی مولکولی مکانیسم گیرنده ها بحث می کند.
حس حساس: سازمان عملکردی و مولکولی تعیین کننده گیرنده های مکانیزه حساس
چکیده
Mechanoreceptors های پوستی در لایه های مختلف پوست قرار دارند که در آنها طیف گسترده ای از محرک های مکانیکی را شامل می شود، از جمله برس، کشش، ارتعاش و فشار مضر. این انواع محرک ها با مجموعه ای متنوع از mehran-receptors های تخصصی است که به شیوه ای خاص به تغییر شکل پوستی پاسخ می دهند و این محرک ها را به ساختارهای مغز بالا می برند. مطالعات در سراسر مكانورزپتورها و انتهای عصب حسی ژنتیکی قابل تشخیص هستند كه مكانیزم لمس احساس را پیدا می كنند. کار در این زمینه، محققان را با درک کامل تر از سازمان مدار که بر پایه درک لمس است، ارائه می دهد. کانال های یون های نوین به عنوان کاندیداهای مولکول های انتقال تبدیل شده اند و خواص جریان های مجهز به مکانیکی، درک ما از مکانیزم های سازگاری با محرک های لمسی را بهبود بخشید. این بررسی پیشرفت هایی را در تعیین ویژگی های عملکردی مکانیسم های گیرنده در پوست های مضر و چشمی و کانال های یونی نشان می دهد که ورودی های مکانیکی را شکل می دهند و سازگاری مکانیس گیر را شکل می دهند.
لمس تشخیص محرک مکانیکی بر روی پوست، از جمله محرک های مکانیکی بی ضرر و مضر است. این یک ضرورت حیاتی برای بقا و توسعه پستانداران و انسان است. تماس با اشیاء جامد و مایعات با پوست اطلاعات لازم را به سیستم عصبی مرکزی می دهد که اجازه می دهد اکتشاف و شناسایی محیط زیست و شروع حرکات حرکتی و حرکت برنامه ریزی شود. لمس نیز برای کارآموزی، ارتباطات اجتماعی و جنسیت بسیار مهم است. حس لمس حس کمترین آسیب پذیر است، اگرچه می توان آن را در بسیاری از شرایط پاتولوژیک (هیپرتانسی، هیپوستزی) تحریف کرد. 1-3
پاسخ های لمسی شامل کدگذاری دقیق اطلاعات مکانیکی است. گیرنده های مکانیکی پوستی در لایه های مختلف پوست محلی هستند که طیف وسیعی از محرک های مکانیکی از جمله برس سبک ، کشش ، لرزش ، انحراف مو و فشار مضر را تشخیص می دهند. این محرک های متنوع با آرایه متنوعی از گیرنده های مکانیکی تخصصی مطابقت دارد که به شکل خاصی به تغییر شکل پوستی پاسخ می دهند و این محرک ها را به ساختارهای بالاتر مغز منتقل می کنند. نورون های حسی حسی پوست به دو گروه تقسیم می شوند: گیرنده های مکانیکی آستانه کم (LTMR) که به فشار خوش خیم واکنش نشان می دهند و گیرنده های مکانیکی آستانه بالا (HTMR) که به تحریک مکانیکی مضر پاسخ می دهند. بدن سلولهای LTMR و HTMR در گانگلیونهای ریشه پشتی (DRG) و گانگلیونهای حسی جمجمه (گانگلیون سه قلو) قرار دارند. الیاف عصبی مرتبط با LTMR ها و HTMR ها بر اساس سرعت هدایت عملکرد بالقوه آنها به عنوان الیاف A - - A ، A - یا C طبقه بندی می شوند. الیاف C فاقد میلین هستند و کمترین سرعت رسانایی را دارند (m 2 m / s) ، در حالی که A؟ و الف؟ الیاف به آرامی و به شدت میلین می شوند ، به ترتیب سرعت هدایت متوسط (~ 12 m / s) و سریع (20 ~ m / s) دارند. LTMR ها نیز با توجه به میزان سازگاری آنها با محرک مکانیکی پایدار ، به عنوان پاسخ های آهسته یا سریع سازگار (SA- و RA-LTMR) طبقه بندی می شوند. آنها بیشتر با اندامهای انتهایی پوستی که عصب می دهند و محرکهای ترجیحی آنها متمایز می شوند.
توانایی مکانیسپتورها برای شناسایی نشانه های مکانیکی به حضور کانال های یون مکانیکی تبدیل می شود که به سرعت نیروهای مکانیکی را به سیگنال های الکتریکی تبدیل می کند و زمینه گیرنده را حذف می کند. این تخریب موضعی، به نام پتانسیل گیرنده، می تواند پتانسیل های عملیاتی را که به سیستم عصبی مرکزی نفوذ می کند تولید کند. با این حال، خواص مولکول هایی که مکانیک انتقال و سازگاری با نیروهای مکانیکی را متمایز می کنند هنوز مشخص نیست.
در این بررسی ، ما مروری داریم بر خواص گیرنده مکانیکی پستانداران در لمس بی ضرر و مضر در پوست مودار و براق. ما همچنین دانش اخیر در مورد خصوصیات جریان مکانیکی دروازه را در تلاش برای توضیح مکانیزم سازگاری گیرنده مکانیکی در نظر می گیریم. سرانجام ، ما پیشرفت های اخیر در شناسایی کانال های یونی و پروتئین های مرتبط مسئول تولید جریان های مکانیکی را بررسی می کنیم.
لمس بی امان
فولیکول مو مرتبط با LTMRs
فولیکول های مو نشان دهنده اندام های کوچک تولید کننده شفت مو است که لمس نور را تشخیص می دهند. الیاف مرتبط با فولیکول های مو به حرکت مو و جهت آن با شلیک قطارهای پتانسیل عمل در هنگام شروع و حذف محرک ها پاسخ می دهند. آنها گیرنده ها را به سرعت در حال انطباق می کنند.
گربه و خرگوش در کت گربه و خرگوش ، فولیکول های مو را می توان به سه نوع فولیکول مو ، داون مو ، محافظ مو و تیلوتریش تقسیم کرد. موهای داون (زیر مو ، پشم ، ولوس) 4 پرشمارترین ، کوتاهترین و مرغوب ترین موهای کت هستند. آنها موج دار ، بی رنگ هستند و در گروه های دو تا چهار تار مو از سوراخ مشترک پوست ظاهر می شوند. موهای گارد (مونوتریش ، شنیدن بیش از حد ، موهای زائد) 4 کمی خمیده ، رنگی یا غیر رنگی هستند و به تنهایی از دهان فولیکول های آنها بیرون می آیند. تیلوتریش ها کمترین ، طولانی ترین و ضخیم ترین موها هستند. 5,6،7 آنها رنگدانه یا غیر رنگی هستند ، گاهی اوقات هر دو هستند و به طور جداگانه از یک فولیکول خارج می شوند که توسط حلقه ای از رگ های خونی مویرگی احاطه شده است. فیبرهای حسی تأمین کننده پیاز مو در زیر غده چربی قرار دارند و به A؟ نسبت داده می شوند؟ یا A؟ -LTMR الیاف .XNUMX
در محدوده نزدیک به پایین شانه مو، درست زیر سطح غدد سباسه حلقه انتهایی لانتانولات پلو - Ruffini است. این انتهای حسی عصبی در یک دوره مارپیچ در اطراف شفت مو درون بافت همبند تشکیل می شود که فولیکول مو دارد. در داخل فولیکول مو، همچنین انتهای عصبی آزاد وجود دارد، بعضی از آنها مکانیسم گیرنده را تشکیل می دهند. اغلب، ذره های لمس (نگاه کنید به پوست خشن) اطراف منطقه گردن فولیکول tylotrich اطراف.
خواص انتهای عصب میلین شده در پوست مودار گربه و خرگوش به طور فزاینده ای در دوره 1930-1970 مورد بررسی قرار گرفته است. و خرگوش ، پاسخ های طبقه بندی شده در سه نوع گیرنده مربوط به حرکات موهای داون (گیرنده های نوع D) ، موهای محافظ (گیرنده های نوع G) و موهای تیلوتریش (گیرنده های نوع T) دارند. 1995 همه پاسخ های فیبر عصبی آوران با هم جمع شده اند در گیرنده Adaply Adapting از نوع I (RA I) با مخالفت با گیرنده Pacinian به نام RA II. گیرنده های مکانیکی RA I سرعت محرک مکانیکی را تشخیص می دهند و دارای مرز تیز هستند. آنها تغییرات حرارتی را تشخیص نمی دهند. Burgess و همکاران همچنین یک گیرنده درست به سرعت سازگار را توصیف می کند که به نوازش پوست یا حرکت چندین مو پاسخ بهینه می دهد ، که به تحریک انتهای پیلو-روفینی نسبت داده می شود. هیچ یک از پاسخهای فولیکول مو به فعالیت فیبر C نسبت داده نشد
موش. در پشتی پوست مودار موش، سه نوع عمده از فولیکول های مو شرح داده شده است: کج و معوج (حدود 72٪)، درفش / auchene (حدود 23٪) و گارد یا tylotrich (حدود 5٪) 11-14 زیگزاگ و درفش / فولیکول موهای آچنه موهای شکننده و کوتاهتر مو را تولید می کند و با یک غدد سباسه همراه است. موهای گارد یا تیلوتریچی بلندترین نوع فولیکول مو هستند. آنها با یک لامپ موی بزرگ همراه با دو غده چربی مشخص می شوند. موها محافظ و awl / auchene در یک الگوی تکراری و به طور منظم فاصله قرار می گیرند در حالی که موهای زنجیلی به شدت مناطق پوستی اطراف دو نوع فولیکول موی بزرگ را پر می کنند. 1 (A1، A2 و A3)].
شکل 1. سازماندهی و پیش بینی های مکانیس گیرنده های پوستی. در پوست مودار، برس و لمس نور به طور عمده توسط غشاء اطراف فولیکول های مو تشخیص داده می شود: awl / auchenne (A1)، zigzag (A2) و گارد (A3). موهای شول / اوچن توسط انتهای لانسیلات C-LTMR (A4) ، A؟ -LTMR و A؟ سه بار عصب کشی می شوند به سرعت سازگار-LTMR (A6). فولیکول های مو زیگزاگ شاخه های کوتاه مو هستند و توسط C-LTMR (A4) و A عصب می شوند؟ -LTMR انتهای لانسولات (A5). طولانی ترین فولیکول های محافظ مو توسط A عصب می شوند؟ به سرعت با انتهای لانسیلات طولی-LTMR سازگار می شوند (A6) و با A ارتباط دارند؟ به آهستگی سازگار-LTMR انتهای گنبد لمسی (A7). پیش بینی های مرکزی همه این الیاف در لایه های مجزا ، اما تا حدی همپوشانی شاخ پشتی نخاع خاتمه می یابد (C-LTMR در لایه II ، Aβ -LTMR در لایه III و Aβ -LTMR در لایه IV و V). پیش بینی های LTMR که فولیکول های مو یا موازی مجاور دارند، به شکل یک ستون باریک در شاخ پشتی نخاعی (B1 در خاکستری) تراز می شوند. فقط در پوست مودار، زیرمجموعه ای از الیاف آزاد C به پایان می رسد اپیدرم را مهار می کند و به لمس لذت بخش پاسخ می دهد (A8). این الیاف C-touch به لمس مضر پاسخ نمی دهند و مسیر حرکت آنها هنوز مشخص نیست (B2). با استفاده از چهار نوع LTMR، در پوست چرب، تماس بی ضرر است. مجتمع سلول نوریت مرکل در لایه پایه اپیدرمی (C1) قرار دارد. این گیرنده مکانیکی از یک آرایش بین بسیاری از سلولهای مرکل و یک پایانه عصبی بزرگ شده از یک A تشکیل شده است؟ فیبر سلول های مرکل، مانند فرآیندهای تماس با کراتینوسیت ها (C2)، انگشت خود را نشان می دهند. پایان Ruffini در dermas محلی است. این یک انتهای حسی کپسوله شده باریک شکل سیگار است که به A متصل است فیبر (C3). پیکره مایسنر به A متصل شده است؟ انتهای عصب و در پاپیلهای پوستی واقع شده است. این مکانیسم گیرنده کپسوله شده از سلولهای حمایت کننده بسته بندی شده که به صورت لامال افقی احاطه شده با بافت همبند (C4) تشکیل شده است، تشکیل شده است. Corpuscular Pacinian مکانیسم گیرنده عمیق تر است. یک تک A؟ انتهای عصب فاقد میلین در مرکز این پیکره بزرگ تخم مرغ ساخته شده از لاماهای متحدالمرکز خاتمه می یابد. پیش بینی این الیاف A؟ -LTMR در نخاع به دو شاخه تقسیم می شود. شاخه مرکزی اصلی (B3) در طناب نخاعی در سوناتا یا اسکیزوفرنی (B5) تشکیل می شود که در آن عضلات اولیه اولین سیناپس خود را ایجاد می کنند (B6). نورونهای ثانویه یک قطعه حساسیتی (B7) را ایجاد می کنند تا یک لننوسوس مدیایی ایجاد کنند که از طریق مغز به میانرشت، بویژه در تالاموس، صعود می کند. شعاع ثانویه LTMR در شاخ عقب در لامینا II، IV، V متوقف می شود و با انتقال درد (B4) مواجه می شود. لمس مزاحم توسط پایان عصب آزاد در اپیدرم هر دو مو (A9) و پوست خشن (C7) تشخیص داده می شود. این گیرنده های مکانیکی انتهای A؟ -HTMR و C-HTMR در تماس نزدیک با کراتینوسیت های همسایه (C6) هستند. A؟ -hTMR در لایه I و V ختم می شود. C-HTMR در لایه I و II (B8) خاتمه می یابد. در سطح شاخ پشتی نخاعی، HTMR های اولیه، سیناپس ها را با نورون های ثانویه ایجاد می کنند که از خط میانی عبور می کنند و به ساختار مغز بالاتر در فاکتوریل پیشانی (B9، B10) صعود می کنند.
اخیراً ، جینتی و همکارانش از ترکیبی از برچسب گذاری ژنتیکی مولکولی و رویکرد ردیابی رتروگراد سوماتوتوپی برای تجسم سازماندهی انتهای آکسونی محیطی و مرکزی LTMR ها در موشها استفاده کردند. توسط سه نوع فولیکول مو استخراج شده و از طریق فعالیت های ترکیبات منحصر به فرد فیبرهای A؟ - ، A؟ - و C به شاخ پشت منتقل می شود.
آنها نشان دادند که برچسب گذاری ژنتیکی سلولهای عصبی تیروزین هیدروکسیلاز مثبت (TH +) DRG جمعیتی از سلولهای عصبی حسی غیرپپتیدرژیک و قطر کوچک را مشخص می کند و امکان تجسم انتهای محیطی C-LTMR را در پوست فراهم می کند. با کمال تعجب ، شاخه های آکسونئال C-LTMR های جداگانه پیدا شده و به انتهای طناب های طولی شکل می گیرند که از نزدیک با زیگزاگ (80 end انتهای) و سول / آوشن (20 of انتهای) همراه است ، اما فولیکول های موی تیلوتریش نیست. 1 (A4)]. مدت ها تصور می شد که انتهای لانسولات طولی منحصراً به A؟ -LTMR تعلق دارد و بنابراین غیر منتظره بود که انتهای C-LTMR انتهای لنگول طولی را تشکیل دهند .15 این C-LTMR ها در مقایسه با آهسته و سریع سازگار یک سازگاری متوسط دارند گیرنده های مکانیکی میلین شده [شکل. 2 (C1)].
شکل 2. گیرنده های لمسی در پستانداران: گیرنده های لمسی جلدی به لمس بی ضرر پشتیبانی شده توسط گیرنده های متعدد با آستانه مکانیکی کم (LTMRs) در پوست بدون کرک و مودار و لمس مضر توسط گیرنده آستانه مکانیکی بالا (HTMR) متمایز می شوند. آنها انتهای بدون اعصاب را تشکیل می دهند که به طور عمده در اپیدرم ختم می شوند. (الف) پوست بدون کرک. A1: اجسام مایسنر حرکت پوست و لیز خوردن شی object در دست را تشخیص می دهند. آنها برای تحویل شی و مهارت بسیار مهم هستند. گیرنده ها به سرعت با محرک سازگار می شوند ، به A متصل می شوند؟ الیاف و به ندرت به الیاف C تبدیل می شوند و دارای میدان گیرنده زیادی هستند. A2: اجسام روفینی کشش پوست را تشخیص می دهند و برای تشخیص موقعیت انگشت و تحویل اشیا مهم هستند. گیرنده به آرامی با محرک سازگار می شود و تا زمانی که محرک اعمال شود ، فعالیت را حفظ می کند. گیرنده ها به A متصل هستند؟ الیاف و میدان پذیرایی زیادی دارند. A3: پیکرهای پاکیین در درم عمیق تر هستند و ارتعاش را تشخیص می دهند. گیرنده ها به A متصل هستند؟ الیاف آنها به سرعت با محرک سازگار می شوند و بزرگترین میدان پذیرایی را دارند. (B) پوست کامل. B1: کمپلکسهای سلول مرکل هم در پوست بدون کدر و هم در اطراف مو وجود دارد. آنها به صورت متراکم در دست بیان می شوند و برای درک بافت و بهترین تبعیض بین دو نقطه مهم هستند. آنها مسئول دقت انگشت هستند. گیرنده ها به A متصل هستند؟ الیاف آنها به آرامی با محرک سازگار می شوند و زمینه پذیرایی کوتاهی دارند. B2: HTMR های لمسی مضر با سازگاری بسیار کند با محرک ، یعنی تا زمانی که محرک درد اعمال شود ، فعال هستند. آنها با انتهای عصب آزاد A تشکیل می شوند؟ و فیبرهای C مرتبط با کراتینوسیت ها. (ج) پوست مو دار. C1: فولیکول های مو با انواع مختلف مو ارتباط دارند. در موش موهای محافظ بلندتر و کم بیان شده است ، سول / آوشن اندازه متوسط و زیگزاگ کوچکترین و متراکم ترین مو است. آنها به A متصل هستند؟ الیاف بلکه به A؟ و الیاف C-LTMR برای موهای سولفات / آوشن و زیزاگ. آنها حرکت مو از جمله لمس دلپذیر هنگام نوازش را تشخیص می دهند. آنها به سرعت یا با جنبشی متوسط به محرک سازگار می شوند. C2: انتهای عصب C-touch با یک نوع فرعی از انتهای رشته های C با انتهای آزاد مطابقت دارد که با آستانه مکانیکی کم مشخص می شود. آنها قرار است رمزگذاری کنند برای یک احساس خوشایند ناشی از نوازش. آنها به طور متوسط با محرک سازگار می شوند و زمینه پذیرایی کوتاهی دارند. کانالهای یونی مکانیکی حساس قلمداد شده (MS) بیان شده در گیرنده های مختلف لمسی مطابق با داده های اولیه نشان داده شده و فرضیه موجود تحت ارزیابی را خلاصه می کند.
دومین جمعیت اصلی مشخص شده مربوط به انتهای A؟ -LTMR در فولیکول های Awl / Auchenne و zigzag است تا با فولیکول مو داون که به طور گسترده در گربه و خرگوش مورد مطالعه قرار گرفته مقایسه شود. جینتی و همکارانش نشان دادند که TrkB در زیر مجموعه ای از نورون های DRG با قطر متوسط در سطوح بالا بیان می شود. ضبط های داخل سلولی با استفاده از آماده سازی ex-vivo پوست و عصب الیاف دارای برچسب نشان می دهد که آنها ویژگی های فیزیولوژیکی الیاف قبلاً مورد بررسی در گربه و خرگوش را نشان می دهند: حساسیت مکانیکی نفیس (آستانه Von Frey <0.07 mN) ، پاسخ سریع به محرک های فوق آستانه ، هدایت میانی سرعت (5.8/0.9 15 1/5 متر بر ثانیه) و خوشه های سوما غیر انعطاف پذیر باریک است .XNUMX این A؟ -LTMR ها انتهای طولی لانسولات را تشکیل می دهند که تقریباً با هر پیاز موی زیگزاگ و شول / اوچن تنه مرتبط است [شکل. XNUMX (AXNUMX)].
سرانجام ، آنها نشان دادند که پایان های پیرامونی سازگاری سریع A؟ LTMR ها انتهای طولی متصل به طناب را تشکیل می دهند که با محافظ (یا تیلوتریش) و فولیکول های موی سولف / اوشن مرتبط هستند [شکل 1 (A6)] .15 علاوه بر این ، موهای محافظ نیز با یک مجموعه سلول مرکل در ارتباط هستند که یک گنبد لمسی متصل به A را تشکیل می دهد؟ به آرامی با LTMR سازگار می شود [شکل. 1 (A7)].
به طور خلاصه ، تقریباً تمام فولیکول های موی زیگزاگ توسط هر دو انتهای حلقوی C-LTMR و A؟ -LTMR عصب کشی می شوند. موهای اولی / اوچن توسط A عصب سه برابر می شوند؟ به سرعت با پایان دادن به لانسولات LTMR ، A؟ -LTMR و C-LTMR سازگار می شوند. فولیکول های مو محافظ توسط A عصب می شوند؟ به سرعت با پایان های لانسی طولی-LTMR سازگار می شود و با A تعامل دارد؟ به آهستگی سازگار-LTMR از انتهای گنبد لمسی. بنابراین ، هر فولیکول موی موش ترکیبی منحصر به فرد و ثابت از انتهای LTMR مربوط به اندام های انتهایی مکانیوسحسی مغز و اعصاب متمایز دریافت می کند. با توجه به آرایش تکراری این سه نوع مو ، جینتی و همکارانش پیشنهاد می کنند که پوست مو شامل تکرار تکراری واحد محیطی حاوی ، (1) یک یا دو مو محافظ واقع در مرکز ، (2) ~ 20 surrounding موهای شول / اوشن اطراف و (3 ) ~ 80 موی زیگزاگ متقاطع [شکل. 2 (C1)].
رونده طناب نخاعی. پیش بینی های مرکزی A؟ به سرعت سازگار-LTMR ها ، A؟ -LTMR ها و C-LTMR ها در لایه های متمایز ، اما تا حدودی همپوشانی (II ، III ، IV) شاخ پشتی نخاع ختم می شوند. علاوه بر این ، پایانه های مرکزی LTMR که فولیکول های موی یکسان یا مجاور را در یک واحد LTMR محیطی عصب کشی می کنند ، در یک راستا قرار گرفته و یک ستون باریک LTMR در شاخ پشتی نخاع تشکیل می دهند [شکل. 1 (B1)]. بنابراین ، به نظر می رسد که یک گوه ، یا ستونی از انتهای آوران حسی اولیه حرکتی سازمان یافته در شاخ پشتی نشان دهنده ترازبینی پیش بینی های مرکزی A؟ - ، A؟ - و C-LTMR است که همان واحد محیطی را عصب می دهد و مکانیکی را تشخیص می دهد محرک هایی که بر همان گروه کوچک فولیکول های مو تأثیر می گذارند. بر اساس تعداد موهای محافظ ، شول / اوچن و زیگزاگ تنه و اندام ها و تعداد هر زیرگروه LTMR ، جینتی و همکاران تخمین می زنند که شاخ پشتی ماوس شامل 2,000 تا 4,000 ستون LTMR باشد ، که مطابق با تقریبی تعداد محیط های اطراف است. واحدهای LTMR .15
علاوه بر این ، آکسون های زیرگروه LTMR از نزدیک با یکدیگر در ارتباط هستند ، دارای پیش بینی های درهم تنیده و انتهای لانسولات یکپارچه شده که همان فولیکول مو را عصبی می کند. علاوه بر این ، از آنجا که سه نوع فولیکول مو اشکال ، اندازه ها و ترکیبات سلولی مختلفی از خود نشان می دهند ، احتمالاً دارای خواص تنظیم انحراف یا تنظیم ارتعاشی متفاوتی هستند. این یافته ها با اندازه گیری های کلاسیک مغز و اعصاب فیزیکی در گربه و خرگوش مطابقت دارد که نشان می دهد A؟ RA-LTMR ها و A--LTMR ها می توانند با تغییر شکل انواع مختلف فولیکول مو فعال شوند. 16,17،XNUMX
به این ترتیب، در پوست مودار لمس ترکیبی از (1) تعداد نسبی، توزیع فضایی منحصر به فرد و ویژگی های متمایز مورفولوژیک و انحراف از سه نوع فولیکول های مو است؛ (2) ترکیبی منحصر به فرد از انتهای زیرمطمئن LTMR در ارتباط با هر یک از سه نوع فولیکول مو؛ و (3) حساسیت های متمایز، سرعت هدایت، الگوهای قطار سنبله و خواص سازگاری چهار طبقه اصلی از LTMR مرتبط با فولیکول های مو است که سیستم مکانیسنسوری پوست مو را قادر می سازد استخراج و انتقال به ترکیبات پیچیده از ویژگی های که یک دست زدن به.
LTMRs End-of-Free عصبی
به طور کلی ، انتهای آزاد فیبرهای C در پوست HTMR است ، اما یک زیر جمعیت از الیاف C به لمس مضر پاسخ نمی دهد. این زیرمجموع آوران لمسی فیبر C (CT) نشان دهنده نوع متمایزی از واحدهای گیرنده مکانیکی بدون میلین و آستانه کم است که در پوست مودار اما بدون براق انسان و پستانداران وجود دارد [شکل. 1 (A8)]. 18,19،20,21 CT به طور کلی با درک تحریک لمسی دلپذیر در تماس با بدن مرتبط است. XNUMX،XNUMX
آوران CT به نیروهای تورفتگی در محدوده 0.3 2.5 mN پاسخ می دهند و بنابراین به اندازه بسیاری از A به تغییر شکل پوست حساس هستند؟ آوران ها .19 ویژگی های سازگاری آوران CT در مقایسه با گیرنده های مکانیزه میلین به آرامی و به سرعت سازگار است. زمینه های پذیرای آوران CT انسان تقریباً گرد یا بیضی شکل است. این میدان شامل یك تا نه لكه پاسخگو كوچك است كه در یك منطقه تا 35 mm2.22 توزیع شده است. 50 گیرنده های همولوگ موش در الگویی از وصله های ناپیوسته سازمان یافته اند كه حدود 60-2٪ از ناحیه پوست مو را پوشش می دهد 2 (C23)]. XNUMX
شواهد موجود در مورد بیماران مبتلا به کم خونی ناشی از میلینین نشان می دهد که سیگنالینگ در فیبرهای CT موجب تقویت قشر ساحلی می شود. از آنجا که این سیستم را پشتیبانی می کند فقیر در جنبه های افتراقی لمس است، اما به خوبی مناسب برای رمزگذاری آهسته، لمس ملایم است، CT الیاف در پوست مودار ممکن است بخشی از یک سیستم برای پردازش جنبه های لذت بخش و اجتماعی مربوط به فعال سازی فیبر touch.24 CT همچنین ممکن است نقش مهمی در درد ایفا می کند و اخیرا پیش بینی شده است که التهاب یا تروما ممکن است احساسات انتقال LTMR های C-fiber را از لمس دلپذیر به درد تغییر دهد. 25,26
کدام مسیر سفرهای CT-afferents هنوز شناخته نشده است [شکل. 1 (B2)]، اما کم آستانه لمسی ورودی به سلول های طرح ریزی spinothalamic مستند شده است، 27 وام اعتبار به گزارش از ظریف، کسری مقابل از تشخیص لمسی در بیماران انسانی زیر تخریب این مسیر پس از chordotomy procedures.28
LTMRs در پوست گلی
مجتمع سلول نوریت مرکل و گنبد لمسی. مرکل (1875) اولین کسی بود که توصیف بافتی خوشه های سلولهای اپیدرمی را با هسته های بزرگ لوبوله ای ارائه داد و با فیبرهای عصبی آوران فرضی تماس برقرار کرد. وی تصور كرد كه آنها با لمس كردن آنها Tastzellen (سلولهای لمسی) حس لامسه را تسخیر می كنند. در انسان ، مجتمع های سلول نئوریت مرکل در مناطق حساس به لمس پوست غنی می شوند ، آنها در لایه پایه اپیدرم در انگشتان ، لب ها و اندام های تناسلی یافت می شوند. آنها همچنین در پوست مودار با تراکم کمتری وجود دارند. مجتمع سلول نئوریت مرکل از یک سلول مرکل در موقعیت نزدیک به یک ترمینال عصب بزرگ شده از یک A میلین شده واحد تشکیل شده است. فیبر [شکل 1 (C1)] (مروری در Halata و همكاران) .29 در قسمت اپیدرمی سلول مرکل فرایندهای انگشت مانند گسترش یافته بین كراتینوسیت های همسایه را نشان می دهد [شکل. 1 (C2)]. سلولهای مرکل سلولهای اپیدرمی مشتق شده از کراتینوسیت هستند. 30,31،150 اصطلاح گنبد لمسی برای نامگذاری غلظت زیادی از کمپلکسهای سلول مرکل در پوست مودار قسمت جلوی پنجه گربه معرفی شد. یک گنبد لمسی می تواند حداکثر 32 سلول مرکل داشته باشد که توسط یک A-فیبر و در انسان علاوه بر الیاف A؟ -A ، A؟ و الیاف C نیز به طور منظم وجود داشت. 34-XNUMX
تحریک مجتمع های سلول نئوریت مرکل در پاسخ های نوع I (SA I) با آهستگی سازگار می شود ، که از میدان های پذیرای با مرزهای تیز سرچشمه می گیرند. هیچ تخلیه خود به خودی وجود ندارد. این مجموعه ها به عمق فرورفتگی پوست پاسخ می دهند و بیشترین تفکیک مکانی (0.5 میلی متر) گیرنده های مکانیکی پوست را دارند. آنها تصویر مکانی دقیق از محرکهای لمسی را منتقل می کنند و پیشنهاد می شود مسئول تبعیض شکل و بافت باشند [شکل. 2 (B1)]. موش های فاقد سلول های مرکل در حالی که این کار را با استفاده از سبیل های خود انجام می دهند ، نمی توانند سطوح بافت را با پا تشخیص دهند
این که سلول مرکل ، نورون حسی یا هر دو مکان انتقال مکانیکی هستند هنوز محل بحث است. در موشهای صحرایی ، تخریب سمیت سلولی سلولهای مرکل پاسخ SA I را از بین می برد .36 در موشهایی که سلولهای تحت فشار ژنتیکی سرکوب شده ژنتیکی دارند ، پاسخ SA I که در آماده سازی پوست / عصب در داخل بدن ثبت شده است کاملا ناپدید شد ، نشان می دهد که سلولهای مرکل برای کدگذاری مناسب مرکل مورد نیاز هستند پاسخهای گیرنده .37 با این حال ، تحریک مکانیکی سلولهای مرکل جدا شده در فرهنگ توسط فشار موتور حرکتی جریان مکانیکی را ایجاد نمی کند. 38,39،XNUMX کراتینوسیت ها ممکن است نقش مهمی در عملکرد طبیعی مجموعه سلولهای مغز و اعصاب مرکل داشته باشند. فرآیندهای انگشت مانند سلول مرکل می توانند با تغییر شکل پوست و حرکت سلول اپیدرم حرکت کنند و این ممکن است اولین گام انتقال مکانیکی باشد. واضح است که هنوز شرایط لازم برای مطالعه حساسیت مکانیکی سلولهای مرکل مشخص نشده است.
انتهای Ruffini انتهای روفینی انتهای حسی کپسوله ای نازک سیگار برگ هستند که به A متصل هستند؟ انتهای عصبی انتهای روفینی سیلندرهای بافت همبند کوچکی است که در امتداد رشته های کلاژن پوستی قرار گرفته اند و توسط یک تا سه رشته عصبی میلین شده به قطر 4 6 متر تهیه می شوند. ممکن است حداکثر سه استوانه با جهت گیری متفاوت درم ادغام شده و یک گیرنده ایجاد شود [شکل. 1 (C3)]. از نظر ساختار ، انتهای روفینی شبیه اندام تاندون گلژی است. آنها به طور گسترده در درم بیان می شوند و به عنوان گیرنده های مکانیکی پوستی نوع II (SA II) که به آرامی سازگار هستند ، شناسایی شده اند. در برابر فعالیت عصبی خود به خود ، تخلیه منظم آهسته سازگار با نیروی کم عمود و محرک مکانیکی حفظ می شود یا به طور م effectivelyثرتری توسط کشش پوستی ایجاد می شود. پاسخ SA II از زمینه های بزرگ پذیرایی با مرزهای مبهم نشات می گیرد. گیرنده های روفینی به درک جهت حرکت جسم از طریق الگوی کشش پوست کمک می کنند [شکل. 2 (A2)].
در موش، پاسخ های SA I و SA II را می توان از طریق الکتروفیزیولوژیکی در آماده سازی پوست عصبی ex vivo جدا کرد. 40 Nandasena و همکاران ایمونولوکالیزه کردن آکوپورین 1 (AQP1) در انتهای پریودنتال Ruffini موش های موش صحرایی نشان می دهند که AQP1 درگیر حفظ تعادل اسمزی دندانی لازم برای مکانیک انتقال. 41 پایان دوره پریودنتال Ruffini همچنین بیان کانال ion کانون احتمالی ASIC3.42
مرض مايسنر پیکره های مایسنر در پاپیلاهای پوستی بدون پوست ، به طور عمده در کف دست و کف پا بلکه در لب ها ، در زبان ، در صورت ، در نوک سینه ها و در اندام های تناسلی قرار دارند. از نظر آناتومیک ، آنها از یک انتهای عصبی کپسول دار تشکیل شده اند ، کپسول از سلول های حمایت کننده مسطح ساخته شده است که به صورت لاملهای افقی جاسازی شده در بافت همبند ساخته شده اند. تنها یک فیبر عصبی A وجود دارد؟ آوران هایی که به ازای هر پیکره متصل می شوند [شکل. 1 (C4)]. هر تغییر شکل فیزیکی در پیکره بدن باعث ایجاد یک پتانسیل بالقوه عمل می شود که به سرعت متوقف می شود ، یعنی آنها سریعاً گیرنده ها را سازگار می کنند. وقتی محرک برداشته می شود ، پیکره شکل خود را پیدا می کند و در حین انجام این کار ، پتانسیل های بالقوه دیگری از عمل ایجاد می کند. به دلیل قرارگیری سطحی در درم ، این اجساد به طور انتخابی به حرکت پوست ، تشخیص لمسی و لرزش (20-40 هرتز) پاسخ می دهند. آنها به پوست پویا حساس هستند - به عنوان مثال ، بین پوست و جسمی که تحت آن کار می شود [شکل. 2 (A1)].
قرصهای Pacinian. پیکرهای پاکیین گیرنده های مکانیکی عمیق پوست هستند و حساس ترین گیرنده های پوستی کپسوله شده حرکت پوست هستند. این پیکره های بزرگ تخمک (به طول 1 میلی متر) ساخته شده از لاملا متحدالمرکز از بافت پیوندی رشته ای و فیبروبلاست های پوشیده شده توسط سلول های شوان اصلاح شده تخت در درم عمیق بیان می شوند. 43 در مرکز بدن ، در حفره ای پر از مایع به نام لامپ داخلی ، یک تک A خاتمه می یابد؟ انتهای عصب غیر میلین آور آوران [شکل. 1 (C5)]. آنها دارای یک میدان پذیرایی بزرگ در سطح پوست با یک مرکز حساس هستند. توسعه و عملکرد چندین نوع گیرنده مکانیکی سازگار با سرعت در موشهای جهش یافته c-Maf مختل می شود. به طور خاص ، اجساد پاکیین به شدت آتروفی می شوند
پیکرهای پاکیین در پاسخ به فرورفتگی پوست ، ترشحات عصبی II (RA II) که به سرعت سازگار هستند و قادر به پیروی از فرکانس بالای محرک های ارتعاشی هستند ، سازگاری بسیار سریعی نشان می دهند و از طریق ارتعاشات منتقل شده امکان درک وقایع دور را دارند. آوران ها با شروع فعالیت و تحریک محرک به تورفتگی پایدار با فعالیت گذرا پاسخ می دهند. به آنها آشکارسازهای شتاب نیز گفته می شود زیرا می توانند تغییرات قدرت محرک را تشخیص دهند و اگر سرعت تغییر در محرک تغییر کند (همانطور که در ارتعاشات اتفاق می افتد) ، پاسخ آنها متناسب با این تغییر می شود. پیکره های پاکیین تغییرات فشار فاحش و بیشتر از همه ارتعاشات (45-150 هرتز) را احساس می کنند ، که می توانند حتی سانتی متر از آن را تشخیص دهند [شکل. 300 (A2)].
واکنش تونیک در کپسول دیکاپسولین Pacinian دیده می شود. 46 علاوه بر این، Corpuscles Pacinian بدون عارضه در طول تحرک های دائمی ثابت، بدون تغییر آستانه های مکانیکی یا فرکانس پاسخ، زمانی که سیگنالینگ بین GABA بین glamia lamellate و پایان عصب مسدود می شود. بنابراین، اجزای غیر عصبی مصنوعی Pacinian ممکن است نقش دوگانه در فیلتر کردن محرک مکانیکی و همچنین در تعدیل خواص واکنش نورون حسی داشته باشند.
پیش بینی های طناب نخاعی پیش بینی های A؟ -LTMR در نخاع به دو شاخه تقسیم می شود. شاخه مرکزی اصلی در نخاع در ستون های پشتی یک طرفه به سطح گردن رحم صعود می کند [شکل. 1 (B3)]. شاخه های ثانویه در شاخ پشت در لایه های IV ختم می شوند و به عنوان مثال در انتقال درد اختلال ایجاد می کنند. این ممکن است درد را به عنوان بخشی از کنترل گیت کاهش دهد [شکل. 1 (B4)]. 48
در سطوح رحم، آکسون شاخه اصلی در دو بخش جداگانه می باشد: بخش میانی خطاطی از اطلاعات نیمه پایین بدن (پاها و تنه) حاوی اطلاعات فیزیولوژیکی شکلی است و بخش بیرونی حاوی اطلاعات انتقال تراشه از نیمه بالایی از بدن (بازوی و تنه) [شکل. 1 (B5)].
آوران های لمسی اولیه اولین سیناپس خود را با سلولهای عصبی مرتبه دوم در مدولا ایجاد می کنند ، جایی که فیبرهای هر دستگاه در هسته ای به همین نام سیناپس می شوند: آکسونهای gracile fasciculus در هسته گرانشی و آکسونهای مهره سیناپس در هسته میخ سیناپس می شوند [شکل. 1 (B6)]. سلولهای عصبی دریافت کننده سیناپس ، آورانهای ثانویه را فراهم می کنند و بلافاصله از خط میانی عبور می کنند و یک تراکت در طرف مقابل ساقه مغز ایجاد می کنند. لمنیسکوس داخلی که از طریق ساقه مغز به ایستگاه رله بعدی در مغز میانی ، به ویژه در تالاموس صعود می کند. . 1 (B7)].
مشخصات مولکولی LTMRs. مکانیسم های مولکولی کنترل تنوع اولیه LTMR اخیراً تا حدی روشن شده است. Bourane و همکارانش نشان داده اند که جمعیتهای عصبی بیان کننده گیرنده تیروزین کیناز Ret (Ret) و گیرنده مشترک آن GFR؟ 2 در موشهای جنینی E11 13 DRG به طور انتخابی فاکتور رونویسی Mafa را بیان می کنند. 49,50،2 این نویسندگان نشان می دهند که Mafa / سلولهای عصبی Ret / GFR؟ 1 که قرار است در بدو تولد به سه نوع خاص از LTRM تبدیل شوند: سلولهای عصبی SA51 باعث تحریک مجتمع های سلول مرکل می شوند ، سلولهای عصبی که به سرعت سازگار می شوند و اجسام Meissner را خنثی می کنند و آوران هایی که به سرعت سازگار می شوند (RA I) انتهای لانسولات را در اطراف فولیکول های مو تشکیل می دهند. Ginty و همكارانش همچنین گزارش كردند كه نورونهای DRG كه زود Ret را بیان می كنند به سرعت در حال سازگاری گیرنده های مكانیكی از پیكرهای Meissner ، پیكرهای پاكینین و انتهای لانسولات در اطراف فولیكول های مو هستند. XNUMX آنها مناطق هدف گسسته را در هسته های دانه ای و دانه ای عصب بندی می كنند ، و الگوی خاصی از مكان حسی را نشان می دهند پیش بینی های آکسون نورون در ساقه مغز.
کشف مکانیسم گیرنده های پوست انسان. تکنیک "میکرونوروگرافی" توصیف شده توسط هاگبارت و والبو در سال 1968 برای مطالعه رفتار تخلیه انتهای مکانیکی حساس انسان تأمین کننده عضله ، مفصل و پوست استفاده شده است (برای بررسی Macefield ، 2005 مراجعه کنید). 52,53،20 اکثر میکروارونروگرافی پوست انسان مطالعات فیزیولوژی آوران لمسی در پوست براق دست را مشخص کرده است. ضبط های میکروالکترود از اعصاب میانی و اولنار در افراد انسانی ، احساس لمس ایجاد شده توسط چهار گروه LTMR را نشان داده است: آوران مایسنر به ویژه نسبت به سکته های سبک در پوست حساس هستند و به نیروهای برشی محلی و لغزش های اولیه یا واضح در قسمت پذیرایی واکنش نشان می دهند. آورانهای پسیینی به موقتهای مکانیکی سریع حساس هستند. آورندگان با شدت به دمیدن بیش از میدان پذیرش پاسخ می دهند. یک پیکر پاکین واقع در یک رقم معمولاً به ضربه زدن روی میز پشتیبانی کننده بازو پاسخ می دهد. آوران مرکل به طور مشخص دارای حساسیت دینامیکی بالایی به محرک های تورفتگی اعمال شده در یک منطقه گسسته است و اغلب هنگام تخلیه با تخلیه تخلیه پاسخ می دهد. اگرچه آوران های روفینی به نیروهایی که به طور عادی به پوست وارد می شوند پاسخ می دهند ، اما ویژگی منحصر به فرد آوران های SA II توانایی آنها در پاسخ به کشش جانبی پوست است. سرانجام ، واحدهای موی بازو دارای زمینه های بزرگ تخمدانی یا نامنظم هستند که از چندین لکه حساس تشکیل شده اند و با موهای جداگانه مطابقت دارند (هر آورنده XNUMX ~ مو تأمین می کند).
حساسیت مکانیکی کراتینوسیت ها
هر محرک مکانیکی بر روی پوست باید از طریق کراتینوسیت هایی که اپیدرم را تشکیل می دهند، انتقال یابد. این سلول های همه جا ممکن است علاوه بر نقش های حمایتی یا محافظتی آنها را نیز انجام دهد. به عنوان مثال، کراتینوسیتها ATP، یک مولکول سیگنالینگ حسی حساسیتی را در پاسخ به محرک مکانیکی و اسموتیک ترشح می کنند. 54,55 انتشار ATP باعث افزایش میزان کلسیم داخل سلولی توسط تحریک اتوکرین از گیرنده های پورینرژیک می شود. 55 علاوه بر این، شواهدی وجود دارد که هیپوتونیکیت Rho-kinase مسیر سیگنالینگ و شکل گیری فیبرهای تنش بعدی F-actin نشان می دهد که تغییر شکل مکانیکی کراتینوسیت ها می تواند به صورت مکانیکی با سلول های همسایه مانند سلول های مرجک برای لکه های بی خطر و پایان انتهایی آزاد C به لمس زخم نفوذ کند. 1 (C6)] 56,57
لمسی مضر
گیرنده های مکانیکی آستانه بالا (HTMR) C- و A اپیدرمی هستند؟ انتهای عصبی رایگان آنها با ساختارهای خاصی همراه نیستند و در هر دو پوست مو مشاهده می شوند [شکل. 1 (A9)] و پوست بدون براق [شکل. 1 (C7)]. با این حال ، اصطلاح انتهای عصب آزاد باید محتاطانه در نظر گرفته شود زیرا انتهای عصب همیشه با سلولهای کراتینوسیت یا سلول لانگرانس یا ملانوسیت ها در موقعیت نزدیک قرار دارند. تجزیه و تحلیل فراساختاری انتهای عصب وجود شبکه آندوپلاسمی خشن ، میتوکندری فراوان و وزیکول هسته متراکم را نشان می دهد. غشاهای مجاور سلولهای اپیدرم ضخیم شده و شبیه غشای پس سیناپسی در بافتهای عصبی است. توجه داشته باشید که فعل و انفعالات بین انتهای عصب و سلولهای اپیدرمی ممکن است دو طرفه باشد زیرا سلولهای اپیدرمی ممکن است واسطه هایی را به عنوان ATP ، اینترلوکین (IL6 ، IL10) و برادی کینین آزاد کنند و بالعکس انتهای عصب پپتیدرژیک می تواند پپتیدهایی مانند CGRP یا ماده P را که بر سلولهای اپیدمی تأثیر می گذارد آزاد کند. HTMR ها شامل گیرنده های مکانیکی-مکانیکی هستند که فقط توسط محرک های مکانیکی مضر و گیرنده های چند مدلی تحریک می شوند که همچنین به گرمای مضر و مواد شیمیایی برونزا پاسخ می دهند [شکل. 2 (B2)]. 58
الیاف آوران HTMR در سلولهای عصبی فرافکن در شاخ پشتی نخاع خاتمه می یابند. A -HTMR ها غالباً در لایه های I و V با سلول های عصبی مرتبه دوم تماس می گیرند ، در حالیکه C-HTMR در لایه II خاتمه می یابد [شکل. 1 (B8)]. سلولهای عصبی درد مرتبه دوم به طرف متقابل نخاع منتقل می شوند و در ماده سفید صعود می کنند و سیستم قدامی استخوان را تشکیل می دهند. این نورون ها عمدتا در تالاموس ختم می شوند [شکل. 1 (B9 و B10)].
مکانیسم جریان در نورونهای Somatosensory
مکانیزم های سازگاری آهسته یا سریع مکانیسم های گیرنده هنوز مشخص نشده است. مشخص نیست که تا چه اندازه سازگاری مکانیسپتورها توسط محیط سلولی انتهای عصبی حسی، خصوصیات ذاتی کانال های مکانیکی و خواص کانال های یونی ولتاژ متصل به نورون های حساس (شکل 2) ارائه شده است. با این حال، پیشرفت های اخیر در توصیف جریان های مکانیکی نشان داده است که کلاس های مختلف کانال های حساس مکانیکی در نورون های DRG وجود دارد و ممکن است بعضی از جنبه های سازگاری مکانیزه های گیرنده را توضیح دهد.
ضبط در شرایط آزمایشگاهی در جوندگان نشان داده است که سوما نورونهای DRG ذاتاً مکانیکی و حساس است و بیانگر جریان مکانیکی درب کاتیونی است. 59-64 گادولینیوم و روتنیوم قرمز جریانهای مکانیکی را کاملاً مسدود می کنند ، در حالی که کلسیم و منیزیم خارجی در غلظتهای فیزیولوژیکی و همچنین آمیلورید و بنزامیل ، باعث انسداد جزئی می شود. 60,62,63،1،43 FM1-43 به عنوان یک مسدود کننده ماندگار عمل می کند ، و تزریق FM65-XNUMX به پنجه عقب موش باعث کاهش حساسیت به درد در آزمایش Randall Selitto و افزایش آستانه برداشتن پنجه می شود. XNUMX. موهای فون فری
در پاسخ به تحریک مکانیکی پایدار ، جریانهای حساس به مکانی از طریق بسته شدن کاهش می یابند. بر اساس ثابت های زمان پوسیدگی جریان ، چهار نوع جریان حساس مکانیکی متمایز شده است: جریان های سریع تطبیق پذیر ( 3 6 میلی ثانیه) ، جریان های تطبیقی سریع (15 30 میلی ثانیه،) ، جریان های آهسته سازگار (200 ms 300 میلی ثانیه) ) و جریانهای بسیار آهسته سازگار ((1000 میلی ثانیه) .64 همه این جریانات با بروز متغیری در سلولهای عصبی DRG موش صحرایی وجود دارند که باعث پوسته شدن پوست براق لگن می شود. 64
حساسیت مکانیکی جریانهای حساس به مکانیک را می توان با استفاده از یک سری محرکهای مکانیکی افزایشی تعیین کرد ، که امکان تجزیه و تحلیل جریان تحریک نسبتاً دقیق را فراهم می کند. 66 رابطه جریان تحریک به طور معمول سیگموئید است و حداکثر دامنه جریان با توجه به تعداد کانالهایی که به طور همزمان باز هستند .64,67،63,65 جالب توجه است ، گزارش شده است که جریان مکانیکی حساس به سرعت در حال نمایش آستانه مکانیکی کم و نقطه میانی نیمه فعال است در مقایسه با جریان مکانیکی بسیار آهسته سازگار. XNUMX،XNUMX
نورون های حسینی با فنوتیپ های غیر اختصاصی غیرفعال، به ترتیب به سرعت در حال تطبیق جریان های مکانی حساسیت با آستانه مکانیکی پایین تر بیان می کنند. 60,61,63,64,68 در عوض، به آرامی و بسیار آهسته تطبیق جریان های مکانیزه حساس در بعضی سلول های غیر تصادفی غیر قابل تشخیص گزارش می شود. 64,68 این پیشنهاد را پیشنهاد می کند که این جریان ها ممکن است به آستانه های مکانیکی مختلف در LTMRs و HTMR in vivo دیده می شود. اگرچه این آزمایشات درون آزمایشگاهی باید با احتیاط انجام شود، حمایت از وجود سم در نورونهای DRG مکانیزم های کم و آستانه مکانیکی نیز با تحریک مبتنی بر کشش مبتنی بر کشش نورون های حسی حساس موش صحرایی ارائه شده است. 69 این پارادایم دو جمعیت اصلی نورونهای حساس کششی، که به دامنه کم تحرک پاسخ می دهد و یکی دیگر از آنها که به طور انتخابی به دامنه محرک بالا پاسخ می دهد.
این نتایج پیامدهای مهمی در عین حال احتمالی و مکانیستی دارد: آستانه مکانیسمی از نورونهای حساس ممکن است کمی با سازمان سلولی مکانیسم گیرنده ارتباط داشته باشد، اما ممکن است در خواص کانال های یونی مکانیکی نگه داشته شود.
سازوکارهایی که زمینه ساز حساسیت زدایی جریانهای کاتیونی حساس به مکانیسم در سلول های عصبی DRG موش صحرایی است به تازگی باز شده است. 64,67،70 این مکانیسم از دو مکانیسم همزمان حاصل شده که بر خصوصیات کانال تأثیر می گذارد: سازگاری و غیرفعال سازی. سازگاری برای اولین بار در مطالعات سلول شنوایی مو گزارش شد. می توان آن را از نظر عملیاتی به عنوان ترجمه ای ساده از منحنی فعال سازی کانال مبدل در امتداد محور محرک مکانیکی توصیف کرد. 72-64,67 سازگاری به گیرنده های حسی اجازه می دهد تا حساسیت خود را نسبت به محرک های جدید در حضور محرک موجود حفظ کنند. با این حال ، بخش قابل توجهی از جریانهای حساس به مکانیک در سلولهای عصبی DRG نمی تواند پس از تحریک مکانیکی تهویه مجدداً فعال شود ، که نشان دهنده غیرفعال شدن برخی کانالهای مبدل است. این دو مکانیزم برای کلیه جریانهای حساس به مکانیک شناسایی شده در سلولهای عصبی DRG موش مشترک هستند ، حاکی از آن است که عناصر فیزیکوشیمیایی مرتبط ، سینتیک این کانالها را تعیین می کنند.
در نتیجه ، تعیین خصوصیات جریانهای حساس به درون مکانیکی درون محیطی در تلاش برای شناسایی مکانیزم های انتقال در سطح مولکولی بسیار مهم است. تنوع مشاهده شده در آستانه مکانیکی و سینتیک سازگار جریانهای مکانیکی مختلف در سلولهای عصبی DRG نشان می دهد که ویژگیهای ذاتی کانالهای یونی ممکن است حداقل در بخشی ، آستانه مکانیکی و سینتیک سازگاری گیرنده های مکانیکی توصیف شده در دهه های 1960 را توضیح دهد 80 با استفاده از آماده سازی ex vivo.
پروتئین های ممانعت کننده متضاد
در مقابل، نورونهای somatosensory مونوسانس حساس هستند، اما در مورد هویت مولکول هایی که مکانیک انتقال در پستانداران دارند، کم است. صفحه های ژنتیکی در Drosophila و C. elegans مولکول های مکانیکی انتقال دهنده را شناسایی کرده اند، از جمله خانواده های TRP و Degenerin / Epithelial Na + (Deg / ENaC). 73 تلاش های اخیر برای کشف مبنای مولکولی مکانیک انتقال در پستانداران عمدتا بر همولوگ های این نامزدها متمرکز شده است . علاوه بر این، بسیاری از این نامزدها در مکانیسم گیرنده های پوستی و نورون های سومیوزنسوری حضور دارند (شکل 2).
اسیدهای حساس کانال های یونی
ASIC ها به یک زیرگروه پروتون دار از خانواده کانال Na + degenerin epithelial تعلق دارند .74 سه عضو از خانواده ASIC (ASIC1 ، ASIC2 و ASIC3) در گیرنده های مکانیکی و گیرنده های قلب بیان می شوند. نقش کانالهای ASIC در مطالعات رفتاری با استفاده از موشهایی با حذف هدفمند ژنهای کانال ASIC بررسی شده است. حذف ASIC1 عملکرد گیرنده های مکانیکی پوستی را تغییر نمی دهد بلکه باعث افزایش حساسیت مکانیکی جوندگان می شود. روده را تحت تأثیر قرار می دهد. اختلال ASIC75 باعث کاهش حساسیت مکانیکی آوران احشایی و کاهش پاسخ HTMR های پوستی به محرک های مضر می شود. 2
کانال گیرنده گذرا
خانواده پرستاری TRP به شش زیرفرانسه در پستانداران تقسیم شده است. 78 تقریبا تمام زیرپارامترهای TRP دارای اعضای متصل به مکانیزاسیون در انواع سیستم های سلولی هستند. 79 در نورون های حسی پستانداران، کانال های TRP برای شناخت اطلاعات حرارتی و میانجی گری کردن التهاب نوروژنیک، و تنها دو کانال TRP، TRPV4 و TRPA1، در واکنش لمسی دخالت دارند. اختلال در بیان TRPV4 در موش فقط اثرات منفی بر آستانه های مکانیسونورزی حاد دارد، اما به شدت حساسیت به محرک های مکانیکی مضر را کاهش می دهد. 80,81 TRPV4 یک عامل تعیین کننده در شکل دادن پاسخ نورون های غوطه ور به استرس اسمز و هیپرآلژزیای مکانیکی در طی التهاب است. به نظر می رسد 82,83 TRPA1 نقش مهمی در هیپرالژیزای مكانیك دارد. موشهای کمبود TRPA1 دارای حساسیت به درد هستند. TRPA1 به تحرک محرک های مکانیکی، سرد و شیمیایی در نورون های حسی حساس کمک می کند اما به نظر می رسد که برای انتقال سلول های موی ضروری نیست. 84,85
شواهدی روشن نیست که نشان می دهد کانال های TRP و کانال های ASIC که در پستانداران بیان شده اند از نظر مکانیکی از بین رفته اند. هیچ کدام از این کانال ها به طور heterologous ابراز امید الکتریکی جریان های مکانی حساسیت مشاهده شده در محیط محلی خود را خلاصه نمی کنند. این امر امکان پذیر نیست که کانالهای ASIC و TRP مکانیکاتورها هستند، زیرا عدم اطمینان بودن اینکه آیا یک کانال مکانیکی انتقال ممکن است خارج از بستر سلولی آن عمل کند (بخش را در SLP3 مشاهده کنید).
پروتئین های پیزو
پروتئینهای پیزو که اخیرا به عنوان کاندیدای امیدوارکننده برای پروتئین های مکانیسنسسی توسط کست و همکاران شناخته شده اند. ستون فقرات 86,87 دارای دو عضو پیزو، Piezo 1 و Piezo 2، که قبلا به نام FAM38A و FAM38B شناخته می شوند، که به خوبی در سراسر یوکاریوت چند سلولی محافظت می شوند. Piezo 2 در DRG فراوان است، در حالی که Piezo 1 به سختی قابل تشخیص است. 4 بیان پیزو 88 یا پیزو 1 در سیستم های heterologous ایجاد جریان های مکانیزه حساس، سینتیک غیر فعال شدن Current جی پی یو 2 سریع تر است از Piezo 2 جریانهای وابسته به پیزو مشابه جریانهای مکانی حساس درون درونی، دارای پتانسیل برگشت پذیری در اطراف 1 mV هستند و کاتیونی انتخابی نیستند، با Na +، K +، Ca0 + و Mg2 + همه نفوذ کانال زیر را دارند. به همین ترتیب، جریان وابسته به پیزو توسط پتانسیل غشا تنظیم می شود، با کاهش شدید سینتیک فعلی در پتانسیل های دپولاریزه. 2
پروتئین های پیزو بدون شک پروتئین های مکانیسنزینگ هستند و بسیاری از ویژگی های جریان های مکانی حس حساس را در نورون های حسی به اشتراک می گذارند. درمان از سلول های عصبی DRG کشت مسافرتی المنت دار 2 کوتاه تداخل RNA نسبت از سلول های عصبی با سرعت در حال تطبیق فعلی کاهش یافته و کاهش درصد mechanosensitive حوزه neurones.86 گذرنده در سراسر پروتئین مسافرتی المنت دار واقع اما هیچ نقوش منافذ حاوی آشکار و یا امضا کانال یونی شده شناخته شده است. با این حال، پروتئین Pyzo 1 ماوس پاک شده و به دوشاخه های لیپید نامتقارن تبدیل می شود و لیپوزوم ها کانال های یونی حساس به قرمز روتنیم را تشکیل می دهند. 87 یک گام اساسی در اعتبار دادن مکانیزم انتقال از طریق کانال های پیزو، استفاده از روش های in vivo برای تعیین اهمیت کاربردی در سیگنالینگ لمسی است. اطلاعات در Drosophila که در آن حذف یک عضو پیزو صرفه جویی در واکنش مکانیکی به محرک های خطرناک، بدون تاثیری بر لمس طبیعی، داده شد. 89 اگر چه ساختار آنها هنوز تعیین می شود، این خانواده جدید از پروتئین های مکانیزم حساس، یک موضوع امیدوار کننده برای تحقیقات آینده، فراتر از مرز است احساس لمس برای مثال، یک مطالعه اخیر در مورد بیماران مبتلا به کم خونی (xerocytosis ارثی) نقش پیزو 1 را در حفظ هومیوستاز حجم اریتروسیت نشان می دهد. 90
ترانزیستور کانال مانند (TMC)
یک مطالعه اخیر نشان می دهد که دو پروتئین، TMC1 و TMC2، برای سلول های مویی mechanotransduction.91 ارثی ناشنوایی لازم است با توجه به جهش در ژن TMC1 در انسان و mice.92,93 حضور گزارش شد از این کانال هنوز در سیستم حسی نشان داده شده است نشده است، اما به نظر می رسد که منجر به تحقیق شود.
پروتئین 3 (SLP3) مانند پروتئین Stomatin
علاوه بر کانال های انتقال، برخی از پروتئین های جانبی مرتبط با کانال نیز نقش مهمی در حساسیت لمسی دارند. SLP3 در نورونهای DRG پستانداران بیان شده است. مطالعاتی که با استفاده از موش های جهش بدون SLP3 انجام شد تغییر در مکانیزاسیون و جریان های مکانیکی نشان داده شده است. عملکرد دقیق 94,95 SLP3 هنوز معلوم نیست. ممکن است یک لینک دهنده بین کانال مکانی حساس و میکروتوبولهای زیرزمینی باشد، همانطور که برای C elegans homolog MEC2.96 به تازگی GR پیشنهاد شده است. آزمایشگاه لویون پیشنهاد کرده است که یک تست توسط نورونهای حسی حسگر DRG سنتز شده و کانال یون های مكان حساس را به ماتریس خارج سلولی مرتبط می كند. 97 قطع ارتباط این موج را رفع كننده جریان مكان حساس RA نشان می دهد كه برخی از کانال های یونی تنها حساس به مكان حساس هستند. جریان های مکانیک حساس RA همچنین توسط لامینین-332، یک پروتئین ماتریکس تولید شده توسط کراتینوسیت ها، مهار می شوند، تقویت فرضیه مدولاسیون جریان مکانی حساس با پروتئین های خارج سلولی. 98
زیرشاخه K + کانال
به موازات جریانهای مکانی حساس کاتولیک زدایی کاتالیزور، حضور جریانهای K + مکانی حساس بازتابی می شود. کانال های K + در سلول های ممانعت کننده می توانند در تعادل فعلی گام بردارند و در تعیین آستانه مکانیکی و زمان سازگاری مکانیسم های گیرنده کمک می کنند.
اعضای KCNK متعلق به خانواده کانال K + کانال K2P هستند. 99,100 K2P یک دامنه قابل توجهی از تنظیمات سلولی، فیزیکی و دارویی دارد، از جمله تغییرات pH، گرما، کشش و تغییر شکل غشا. این K2P در پتانسیل غشای استراحت فعال هستند. چندین واحد از KCNK در نورونهای somatosensory ابراز می شوند. کانال های 101 KCNK2 (TREK-1)، KCNK4 (TRAAK) و TREK-2 از کانال های چندگانه است که برای آنها گیتار مکانیکی مستقیم توسط کشش غشاء نشان داده شده است. 102,103
موشهایی که دارای ژن مختل شده KCNK2 هستند ، حساسیت بیشتری نسبت به گرما و محرکهای مکانیکی ملایم نشان می دهند اما یک آستانه طبیعی خروج از فشار مکانیکی مضر را که با استفاده از آزمون Randall Selitto به قسمت عقب وارد می شود ، نشان می دهند. 104 موش کمبود KCNK2 همچنین افزایش بیش فعالی حرارتی و مکانیکی در التهاب را نشان می دهد شرایط موش های ناک اوت KCNK4 نسبت به تحریکات مکانیکی ملایم حساس بودند و این حساسیت بیش از حد با غیرفعال سازی اضافی KCNK2.105 افزایش یافت. افزایش حساسیت مکانیکی این موشک های ناک اوت می تواند بدین معنی باشد که کشش به طور هماهنگ جریان های حساس به مکانیزاسیون و قطبش را به صورت هماهنگ فعال می کند ، همانند عدم تعادل جریانهای دردار ولتاژ دپولاریزه و قطبش مجدد.
KCNK18 (TRESK) یکی از عوامل اصلی در هدایت پس زمینه K + است که پتانسیل غشای استراحت سلولهای عصبی حسی حسی را تنظیم می کند .106 اگرچه مشخص نیست که KCNK18 مستقیماً به تحریک مکانیکی حساس است ، اما ممکن است در واسطه پاسخ به لمس نور نقش داشته باشد ، و همچنین محرکهای مکانیکی دردناک. پیشنهاد می شود KCNK18 و به میزان کمتری KCNK3 هدف مولکولی هیدروکسی -؟ - سانشول باشد ، ترکیبی که در دانه های فلفل Schezuan یافت می شود و گیرنده های لمسی را فعال می کند و احساس سوزن سوزن شدن را در انسان ایجاد می کند. 107,108،XNUMX
KCNQ4 KV کانال K + (Kv7.4) برای تعیین اولویت سرعت و فرکانس زیرمجموعه مکانیسپسپتورهای سریع سازگار در هر دو موش و انسان بسیار مهم است. جهش KCNQ4 در ابتدا با یک شکل ناشنوائی ارثی همراه بود. جالب توجه است که یک مطالعه اخیر، KCNQ4 را در انتهای عصب محیطی از فولیکول مو و مویسنر کولورکتال به سرعت در حال تغذیه پوست قرار می دهد. بر این اساس، از دست دادن عملکرد KCNQ4 منجر به افزایش انتخاب حساسیت مكانورزپتور به لرزش با فركانس پایین می شود. به طور مشخص، افرادی که دچار افت شنوایی دیررس ناشی از جهش های غالب ژن KCNQ4 هستند، عملکرد پیشرفته ای را در تشخیص دامنه های کوچک و لرزش کم نشان می دهند. 109
دکتر الکس جیمنز Insight
لمس به عنوان یکی از پیچیده ترین حواس در بدن انسان محسوب می شود، به ویژه به این دلیل که هیچ ارگان خاصی مسئول آن نیست. در عوض، حس لمس از طریق گیرنده های حسی، به نام mechanoreceptors شناخته می شود که در سراسر پوست یافت می شوند و به فشار مکانیکی یا تحریف می رسند. چهار نوع اصلی از مکانیسپسپتورها در پوست بدون پهن و بدون پوست، پستانداران وجود دارد: سلولهای لایه ای، بافت های لایه ای، انتهای عصب مرکل و سلول های لگن. Mechanoreceptors به منظور تشخیص لمس، به منظور نظارت بر موقعیت عضلات، استخوان ها و مفاصل شناخته شده به عنوان proprioception، و حتی برای تشخیص صداها و حرکت بدن عمل می کند. شناخت مکانیزم های ساختار و عملکرد این مکانیس گیرنده ها، یک عنصر اساسی در استفاده از درمان ها و درمان های درمانی است.
نتیجه
لمس یک حس پیچیده است زیرا نشان دهنده کیفیت های لمسی متفاوت است، یعنی ارتعاش، شکل، بافت، لذت و درد، با عملکرد های مختلف تبعیض آمیز. تا به حال، مکاتبات بین یک عضو لمسی و حس روانشناختی همبستگی و نشانگرهای مولکولی خاص کلاس فقط در حال ظهور است. توسعه تست های جوندگان متناسب با تنوع رفتار لمسی در حال حاضر برای تسهیل شناسایی ژنوم در آینده لازم است. استفاده از موش هایی که دارای زیرمجموعه های خاصی از انواع حساسیت های حساس نیستند، شناسایی گیرنده های مکانیکی و فیبرهای حساس حساس را با روش خاص لمس مشخص می کند. جالب توجه است که یک مقاله اخیر، مسئله مهمی از ژنتیک ویژگی های مکانیسنسوری در انسان را باز می کند و نشان می دهد که جهش تک ژنی می تواند حساسیت لمسی را تحت تأثیر قرار دهد. 110 این امر را تأکید می کند که پاتوفیزیولوژی کسری لمسی انسان در بخش بزرگی ناشناخته است و مطمئنا پیشرفت را با شناسایی دقیق زیر مجموعه نورون های حسی مرتبط با یک روش لمس یا کمبود لمسی.
در عوض، پیشرفت در تعریف خواص بیوفیزیکی جریانهای مکانیکی صورت گرفته است. 64 توسعه تکنیک های جدید در سال های اخیر، اجازه نظارت بر تغییرات تنش غشایی، در حالی که ضبط مکانیکی جریان، روش تجربی ارزشمند برای توصیف جریان های مکانیزه حساس با سازگاری سریع، متوسط و آهسته (بررسی شده در Delmas و همکاران) .66,111 آینده خواهد بود که تعیین نقش خواص جاری در مکانیزم های سازگاری مکانیسم گیرنده های متنوع عملکرد و سهم از جریان های K + مکانیک حساس به تحریک پذیری LTMRs و HTMRs.
طبیعت مولکولی جریان های مکانیکی در پستانداران نیز موضوع پژوهشی امیدوار کننده آینده است. تحقیقات آینده در دو دیدگاه پیشرفت می کند، ابتدا تعیین نقش مولکول های جانبی که کانال های آن را به سلول های سیتواسکل منتقل می کند و مورد نیاز برای تخصیص یا تنظیم مکانی حساسیت کانال های یونی مانند خانواده های TRP و ASIC / EnaC است. دوم، برای بررسی منطقه بزرگ و امیدوار کننده از کانال های پیزو با پاسخ دادن به سوالات کلیدی نسبت به مکانیزم نفوذ و جابجایی، زیر مجموعه نورون های حساس و حالت های لمسی مربوط به پیزو و نقش پیزو در سلول های غیر عصبی مرتبط با مکانیزاسیون
حس لامسه ، در مقایسه با حس بینایی ، چشایی ، صدا و بویایی ، که از اندام های خاصی برای پردازش این احساسات استفاده می کنند ، می تواند در سراسر بدن از طریق گیرنده های ریز شناخته شده به عنوان گیرنده های مکانیکی ایجاد شود. انواع مختلف گیرنده های مکانیکی را می توان در لایه های مختلف پوست یافت ، جایی که آنها می توانند طیف گسترده ای از تحریکات مکانیکی را تشخیص دهند. مقاله فوق نکات برجسته خاصی را نشان می دهد که پیشرفت مکانیسم های ساختاری و عملکردی گیرنده های مکانیکی مرتبط با حس لامسه را نشان می دهد. اطلاعات ارجاع شده از مرکز ملی اطلاعات بیوتکنولوژی (NCBI). دامنه اطلاعات ما محدود به عمل جراحی و همچنین آسیب های ستون فقرات و شرایط است. برای بحث در مورد موضوع ، لطفاً از دکتر جیمنز سوال کنید یا با ما تماس بگیرید915-850-0900.
دکتر الکس جیمنز سرپرستی می کند
موارد اضافی: درد پشت
درد پشت یکی از شایع ترین علل ناتوانی و از دست رفته روز در کار در سراسر جهان است. به عنوان حقيقت، درد پشتي به عنوان دومين شايعترين دلايل بازديد پزشکي، نسبت به عفونت فوقاني تنفسي کمتر است. تقریبا 80 درصد از جمعیت حداقل یک بار در طول زندگی خود نوعی کمر درد را تجربه خواهند کرد. ستون فقرات یک ساختار پیچیده ای است که از استخوان ها، مفاصل، لیگامان ها و ماهیچه ها در میان دیگر بافت های نرم تشکیل شده است. به همین دلیل، آسیب و / یا شرایط شدید، مانند دیسک های فتق دیسک، در نهایت می تواند منجر به علائم درد پشت شود. آسیب های ورزشی یا آسیب های ناشی از تصادفات خودرو اغلب اغلب علت درد پشت هستند، اما گاهی اوقات ساده ترین حرکات می توانند نتایج دردناکی داشته باشند. خوشبختانه، گزینه های درمان جایگزین، مانند مراقبت از کیهان پراکسی، می تواند به کاهش درد در استفاده از تنظیمات ستون فقرات و دستکاری دست کمک کند و در نهایت بهبود تسکین درد را کاهش دهد.
1. موریواکی ک، یوگ او. ویژگیهای توپوگرافی ناهنجاریهای پوستی لمسی و هیپوآستتیک در درد مزمن.درد. 1999.81:1�6. doi: 10.1016/S0304-3959(98)00257-7.�[گروه][صلیب نماینده]
2. Shim B، Kim DW، Kim BH، Nam TS، Leem JW، Chung JM. حساسسازی مکانیکی و حرارتی گیرندههای درد پوستی در موشهای صحرایی مبتلا به نوروپاتی محیطی تجربی.علوم اعصاب. 2005.132: 193. doi: 201/j.neuroscience.10.1016.[گروه]�[صلیب نماینده]
3. Kleggetveit IP، Jórum E. اختلال عملکرد فیبر بزرگ و کوچک در آسیبهای عصبی محیطی با یا بدون درد خود به خود.J Pain. 2010.11: 1305. doi: 10/j.jpain.10.1016.[گروه]�[صلیب نماینده]
4. Noback CR. مورفولوژی و فیلوژنی مو...Ann NY Acad Sci. 1951.53:476�92. doi: 10.1111/j.1749-6632.1951.tb31950.x.�[گروه]�[صلیب نماینده]
5. ما سترایل. فولیکول های آتیپیک محافظ مو در پوست خرگوش.طبیعت . 1958.181:1604�5. doi: 10.1038/1811604a0.�[گروه]�[صلیب نماینده]
6. ما سترایل. مورفولوژی فولیکول های تایلوتریک در پوست خرگوش.من جی آنات.�1961.109: 1. doi: 13/aja.10.1002.�[گروه]�[صلیب نماینده]
9. Brown AG، Iggo A. مطالعه کمی گیرنده های پوستی و فیبرهای آوران در گربه و خرگوش.J Physiol. 1967.193: 707 33. [PMC رایگان مقاله]�[گروه]
10. Burgess PR، Petit D، Warren RM. انواع گیرنده در پوست مودار گربه که توسط فیبرهای میلین دار تامین می شود.J Neurophysiol. 1968.31: 833 48. [گروه]
11. Driskell RR، Giangreco A، Jensen KB، Mulder KW، Watt FM. سلول های پاپیلای پوستی Sox2 مثبت نوع فولیکول مو را در اپیدرم پستانداران مشخص می کنند.توسعه.�2009.136: 2815. doi: 23/dev.10.1242.�[PMC رایگان مقاله]�[گروه]�[صلیب نماینده]
12. حسین م. الگوی کلی آرایش فولیکول های مو در موش و موش.جی آنات.�1971.109: 307 16. [PMC رایگان مقاله]�[گروه]
13. Vielkind U، هاردی MH. تغییر الگوهای مولکول های چسبندگی سلولی در طول رشد فولیکول موی موش 2. مورفوژنز فولیکول در موتانت های مو، تابی و پرزدار.آکتا آنات (بازل)�1996.157: 183 94. doi: 10.1159 / 000147880. [گروه]�[صلیب نماینده]
14. Hardy MH، Vielkind U. تغییر الگوهای مولکول های چسبندگی سلولی در طول رشد فولیکول موی پلاژ موش. 1. مورفوژنز فولیکول در موش های نوع وحشی.آکتا آنات (بازل)�1996.157: 169 82. doi: 10.1159 / 000147879. [گروه]�[صلیب نماینده]
15. Li L، Rutlin M، Abraira VE، Cassidy C، Kus L، Gong S، و همکاران. سازماندهی عملکردی نورون های حسی مکانیکی پوستی با آستانه پایین...سلول. 2011.147: 1615. doi: 27/j.cell.10.1016.[PMC رایگان مقاله]�[گروه]�[صلیب نماینده]
16. Brown AG، Iggo A. مطالعه کمی گیرنده های پوستی و فیبرهای آوران در گربه و خرگوش.J Physiol. 1967.193: 707 33. [PMC رایگان مقاله]�[گروه]
17. Burgess PR، Petit D، Warren RM. انواع گیرنده در پوست مودار گربه که توسط فیبرهای میلین دار تامین می شود.J Neurophysiol. 1968.31: 833 48. [گروه]
18. Vallbo A، Olausson H، Wessberg J، Norrsell U. سیستمی از آوران های بدون میلین برای دریافت مکانیکی بی ضرر در پوست انسان.Brain Res. 1993.628:301�4. doi: 10.1016/0006-8993(93)90968-S.[گروه]�[صلیب نماینده]
19. Vallbo AB، Olausson H، Wessberg J. آورانهای بدون میلین سیستم دومی را تشکیل میدهند که محرکهای لمسی پوست مودار انسان را کدگذاری میکند.J Neurophysiol. 1999.81: 2753 63. [گروه]
22. وسبرگ جی، اولاسون اچ، فرنسترم KW، والبو AB. ویژگی های میدان پذیری آوران های لمسی بدون میلین در پوست انسان.J Neurophysiol. 2003.89: 1567 75. doi: 10.1152/jn.00256.2002.�[گروه][صلیب نماینده]
23. Liu Q، Vrontou S، Rice FL، Zylka MJ، Dong X، Anderson DJ. تجسم ژنتیکی مولکولی زیرمجموعه نادری از نورونهای حسی بدون میلین که ممکن است لمس ملایم را تشخیص دهد.Nat Neurosci. 2007.10: 946. doi: 8/nn10.1038.�[گروه]�[صلیب نماینده]
24. اولاسون اچ، لامار وای، بک لوند اچ، مورین سی، والین بی جی، استارک جی، و همکاران. آوران های لمسی بدون میلین، سیگنالی را به سمت قشر منزوی لمس می کنندNat Neurosci. 2002.5: 900 4. doi: 10.1038/nn896.[گروه]�[صلیب نماینده]
28. سفید JC، شیرین WH. اثربخشی کوردوتومی در درد فانتوم پس از قطع عضوAMA Arch Neurol Psychiatry.�1952.67: 315 22. [گروه]
29. Halata Z، Grim M، Bauman KI. فریدریش زیگموند مرکل و "سلول مرکل" او، مورفولوژی، رشد و فیزیولوژی: بررسی و نتایج جدید.Anat Rec A Discov Mol Cell Evol Biol.�2003.271: 225. doi: 39/ar.a.10.1002.�[گروه]�[صلیب نماینده]
30. Morrison KM، Miesegaes GR، Lumpkin EA، Maricich SM. سلول های مرکل پستانداران از دودمان اپیدرمی منشاء می گیرند.Dev Biol.�2009.336: 76. doi: 83/j.ydbio.10.1016.[PMC رایگان مقاله]�[گروه]�[صلیب نماینده]
31. Van Keymeulen A، Mascre G، Youseff KK، Harel I، Michaux C، De Geest N، و همکاران. اجداد اپیدرمی در طول رشد جنینی و هموستاز بزرگسالان باعث ایجاد سلول های مرکل می شوند.J Cell Biol.�2009.187: 91. doi: 100/jcb.10.1083.�[PMC رایگان مقاله]�[گروه]�[صلیب نماینده]
32. Ebara S، Kumamoto K، Baumann KI، Halata Z. تجزیه و تحلیل سه بعدی گنبدهای لمسی در پوست مودار پنجه گربه، بسترهای مورفولوژیکی را برای پردازش حسی پیچیده نشان می دهد.Neurosci Res.�2008.61: 159. doi: 71/j.neures.10.1016.[گروه]�[صلیب نماینده]
33. کمپلکسهای Guinard D، Usson Y، Guillermet C، Saxod R. Merkel از پوست دیجیتال انسان: تصویربرداری سهبعدی با میکروسکوپ لیزری کانفوکال و ایمونوفلورسانس مضاعف.J Comp Neurol.�1998.398:98�104. doi: 10.1002/(SICI)1096-9861(19980817)398:1<98::AID-CNE6>3.0.CO;2-4.�[گروه][صلیب نماینده]
34. Reinisch CM, Tschachler E. گنبد لمسی در پوست انسان توسط انواع مختلف رشتههای عصبی تامین میشود.آن نورول.2005.58: 88. doi: 95/ana.10.1002.�[گروه]�[صلیب نماینده]
35. Maricich SM، Morrison KM، Mathes EL، Brewer BM. جوندگان برای وظایف تشخیص بافت به سلول های مرکل تکیه می کنند.J Neurosci. 2012.32:3296�300. doi: 10.1523/JNEUROSCI.5307-11.2012.[PMC رایگان مقاله]�[گروه]�[صلیب نماینده]
36. Ikeda I، Yamashita Y، Ono T، Ogawa H. تخریب انتخابی فوتوتوکسیک سلولهای مرکل موشهای صحرایی، پاسخهای واحدهای گیرنده مکانیکی نوع I را که به آرامی تطبیق مییابند، لغو میکند.J Physiol. 1994.479: 247 56. [PMC رایگان مقاله][گروه]
37. Maricich SM، Wellnitz SA، Nelson AM، Lesniak DR، Gerling GJ، Lumpkin EA، و همکاران. سلول های مرکل برای پاسخ های لمس نور ضروری هستندعلم. 2009.324: 1580 �2. doi: 10.1126/science.1172890.[PMC رایگان مقاله]�[گروه]�[صلیب نماینده]
38. دیاموند جی، هلمز ام، پرستار کالیفرنیا. آیا سیناپس های متقابل سلول-نوریت مرکل در شروع پاسخ های لمسی در پوست سمندر نقش دارند؟J Physiol. 1986.376: 101 20. [PMC رایگان مقاله][گروه]
39. Yamashita Y، Akaike N، Wakamori M، Ikeda I، Ogawa H. جریانهای وابسته به ولتاژ در سلولهای مجرد مرکل از موشها.J Physiol. 1992.450: 143 62. [PMC رایگان مقاله]�[گروه]
40. Wellnitz SA، Lesniak DR، Gerling GJ، Lumpkin EA. منظم بودن شلیک مداوم دو گروه از گیرنده های لمسی را که به آرامی تطبیق می یابند در پوست مودار موش نشان می دهد.J Neurophysiol. 2010.103: 3378 88. doi: 10.1152/jn.00810.2009.�[PMC رایگان مقاله]�[گروه]�[صلیب نماینده]
41. Nandasena BG، سوزوکی A، Aita M، Kawano Y، Nozawa-Inoue K، Maeda T. ایمونولوژیک سازی آکواپورین-1 در انتهای روفینی مکانیکی در رباط پریودنتال.Brain Res. 2007.1157: 32. doi: 40/j.brainres.10.1016.[گروه]�[صلیب نماینده]
42. رحمان اف، هارادا اف، سایتو آی، سوزوکی آ، کاوانو وای، ایزومی کی، و همکاران. تشخیص کانال یونی حساس به اسید 3 (ASIC3) در انتهای روفینی پریودنتال دندانهای ثنایای موش.Neurosci Lett. 2011.488: 173. doi: 7/j.neulet.10.1016.[گروه]�[صلیب نماینده]
43. جانسون KO. نقش و عملکرد گیرنده های مکانیکی پوستی.Curr Opin Neurobiol.�2001.11:455�61. doi: 10.1016/S0959-4388(00)00234-8.�[Review]�[گروه]�[صلیب نماینده]
44. Wende H، Lechner SG، Cheret C، Bourane S، Kolanczyk ME، Pattyn A، و همکاران. فاکتور رونویسی c-Maf رشد و عملکرد گیرنده لمسی را کنترل می کندعلم. 2012.335:1373. doi: 6/science.10.1126.�[گروه]�[صلیب نماینده]
49. Bourane S، Garces A، Venteo S، Pattyn A، Hubert T، Fichard A، و همکاران. زیرگروه های گیرنده مکانیکی آستانه پایین به طور انتخابی MafA را بیان می کنند و با سیگنالینگ Ret مشخص می شوند.نورون. 2009.64: 857 70. doi: 10.1016/j.neuron.2009.12.004.[گروه]�[صلیب نماینده]
50. Kramer I، Sigrist M، de Nooij JC، Taniuchi I، Jessell TM، Arber S. نقشی برای سیگنال دهی فاکتور رونویسی Runx در تنوع نورون حسی گانگلیون ریشه پشتی.نورون. 2006.49: 379 93. doi: 10.1016/j.neuron.2006.01.008.[گروه]�[صلیب نماینده]
51. Luo W، Enomoto H، Rice FL، Milbrandt J، Ginty DD. شناسایی مولکولی گیرنده های مکانیکی سریع تطبیق پذیر و وابستگی تکوینی آنها به سیگنالینگ ret.نورون. 2009.64: 841 56. doi: 10.1016/j.neuron.2009.11.003.[PMC رایگان مقاله]�[گروه]�[صلیب نماینده]
52. Vallbo AB، Hagbarth KE. فعالیت از گیرنده های مکانیکی پوست به صورت از راه پوست در افراد بیدار ثبت شده استاکسپ نورول.�1968.21:270�89. doi: 10.1016/0014-4886(68)90041-1.�[گروه]�[صلیب نماینده]
53. Macefield VG. ویژگی های فیزیولوژیکی گیرنده های مکانیکی آستانه پایین در مفاصل، ماهیچه ها و پوست در افراد انسانی.Clin Exp Pharmacol Physiol.�2005.32:135�44. doi: 10.1111/j.1440-1681.2005.04143.x.�[Review]�[گروه]�[صلیب نماینده]
54. Koizumi S، Fujishita K، Inoue K، Shigemoto-Mogami Y، Tsuda M، Inoue K. امواج Ca2+ در کراتینوسیت ها به نورون های حسی منتقل می شوند: دخالت ATP خارج سلولی و فعال شدن گیرنده P2Y2.بیوشیم جی2004.380: 329. doi: 38/BJ10.1042.�[PMC رایگان مقاله]�[گروه][صلیب نماینده]
55. Azorin N، Raoux M، Rodat-Despoix L، Merrot T، Delmas P، Crest M. سیگنال دهی ATP برای پاسخ کراتینوسیت های انسانی به تحریک مکانیکی توسط شوک هیپواسموتیک بسیار مهم است.اکسپ درماتول.�2011.20:401�7. doi: 10.1111/j.1600-0625.2010.01219.x.�[گروه]�[صلیب نماینده]
56. Amano M، Fukata Y، Kaibuchi K. تنظیم و عملکرد کیناز Rho مرتبط.Exp Cell Res.�2000.261: 44. doi: 51/excr.10.1006.�[بازبینی]�[گروه]�[صلیب نماینده]
57. Koyama T، Oike M، Ito Y. دخالت روکیناز و تیروزین کیناز در انتشار ATP ناشی از استرس هیپوتونیک در سلولهای اندوتلیال آئورت گاوی.J Physiol. 2001.532:759�69. doi: 10.1111/j.1469-7793.2001.0759e.x.�[PMC رایگان مقاله]�[گروه]�[صلیب نماینده]
58. پرل ای آر. گیرنده های پوستی چند وجهی: ویژگی ها و شکل پذیری.Prog Brain Res.�1996.113:21�37. doi: 10.1016/S0079-6123(08)61079-1.�[Review]�[گروه]�[صلیب نماینده]
59. McCarter GC، Reichling DB، Levine JD. انتقال مکانیکی توسط نورون های گانگلیونی ریشه پشتی موش در شرایط آزمایشگاهی.Neurosci Lett. 1999.273:179�82. doi: 10.1016/S0304-3940(99)00665-5.�[گروه]�[صلیب نماینده]
60. Drew LJ، Wood JN، Cesare P. خصوصیات مکانیکی حساس نورونهای حسی حساس به کپسایسین و غیر حساس.J Neurosci. 2002.22: RC228. [گروه]
61. Drew LJ، Rohrer DK، Price MP، Blaver KE، Cockayne DA، Cesare P، و همکاران. کانال های یونی حساس به اسید ASIC2 و ASIC3 به جریان های مکانیکی فعال در نورون های حسی پستانداران کمک نمی کنند.J Physiol. 2004.556: 691 710. doi: 10.1113/jphysiol.2003.058693.[PMC رایگان مقاله]�[گروه]�[صلیب نماینده]
62. مک کارتر جی سی، لوین جی دی. اساس یونی جریان مکانیکی در نورونهای گانگلیون ریشه پشتی موش بالغ.مول درد. 2006.2:28. doi: 10.1186/1744-8069-2-28.�[PMC رایگان مقاله]�[گروه]�[صلیب نماینده]
63. Coste B، Crest M، Delmas P. تشریح فارماکولوژیک و توزیع NaN/Nav1.9، جریانهای Ca2+ نوع T و جریانهای کاتیونی فعال شده مکانیکی در جمعیتهای مختلف نورونهای DRG.جی ژن فیزیول.�2007.129:57. doi: 77/jgp.10.1085.�[PMC رایگان مقاله]�[گروه]�[صلیب نماینده]
64. Hao J، Delmas P. مکانیسمهای چندگانه حساسیت زدایی کانالهای مبدل مکانیکی، شلیک نورونهای حسی مکانیکی را شکل میدهند.J Neurosci. 2010.30:13384�95. doi: 10.1523/JNEUROSCI.2926-10.2010.[گروه]�[صلیب نماینده]
65. درو ال جی، وود جی ان. FM1-43 مسدودکننده کانال های یونی حساس به مکانیسم در نورون های حسی است و واکنش های رفتاری به محرک های مکانیکی را مهار می کند.مول درد. 2007.3:1. doi: 10.1186/1744-8069-3-1.�[PMC رایگان مقاله]�[گروه]�[صلیب نماینده]
66. Hao J, Delmas P. ضبط جریانهای حساس به مکانیکی با استفاده از محرک مکانیکی پیزوالکتریک.Nat Protoc.�2011.6:979×90. doi: 10.1038/nprot.2011.343.�[گروه]�[صلیب نماینده]
68. هو جی ، لوین جی. جریانهای حساس به مکانیسم در نوریتهای سلولهای عصبی حسی موش کشت شدهJ Physiol. 2006.577: 815 28. doi: 10.1113/jphysiol.2006.117648.[PMC رایگان مقاله]�[گروه]�[صلیب نماینده]
69. Bhattacharya MR ، Bautista DM ، Wu K ، Haeberle H ، Lumpkin EA ، Julius D. کشش شعاعی جمعیت متمایز نورونهای حسی حسی پستانداران مکانیکی را نشان می دهد.Proc Natl Acad Sci US A. 2008.105:20015. doi: 20/pnas.10.1073.�[PMC رایگان مقاله]�[گروه]�[صلیب نماینده]
70. Crawford AC، Evans MG، Fettiplace R. فعالسازی و سازگاری جریانهای مبدل در سلولهای موی لاکپشت.J Physiol. 1989.419: 405 34. [PMC رایگان مقاله]�[گروه]
71. Ricci AJ، Wu YC، Fettiplace R. بافر کلسیم درونزا و دوره زمانی سازگاری مبدل در سلولهای موی شنوایی.J Neurosci. 1998.18: 8261 77. [گروه]
85. Kwan KY، Allchorne AJ، Vollrath MA، Christensen AP، Zhang DS، Woolf CJ، و همکاران. TRPA1 به درد سرد، مکانیکی و شیمیایی کمک می کند اما برای انتقال سلول های مو ضروری نیست.نورون. 2006.50: 277 89. doi: 10.1016/j.neuron.2006.03.042.[گروه]�[صلیب نماینده]
88. Bae C، Sachs F، Gottlieb PA. کانال یونی حساس به مکانیک Piezo1 توسط پپتید GsMTx4 مهار می شود.بیوشیمی.2011.50: 6295 300. doi: 10.1021/bi200770q.�[PMC رایگان مقاله]�[گروه][صلیب نماینده]
89. Kim SE، Coste B، Chadha A، Cook B، Patapoutian A. نقش مگس سرکه در دردهای مکانیکی.طبیعت . 2012.483: 209. doi: 12/nature10.1038.�[PMC رایگان مقاله]�[گروه][صلیب نماینده]
90. Zarychanski R، Schulz VP، Houston BL، Maksimova Y، Houston DS، Smith B، و همکاران. جهش در پروتئین انتقال مکانیکی PIEZO1 با خشکی سلولی ارثی مرتبط است.خون.2012.120:1908�15. doi: 10.1182/blood-2012-04-422253.�[PMC رایگان مقاله]�[گروه]�[صلیب نماینده]
91. کاواشیما Y، گلوک GS، Kurima K، Labay V، Lelli A، Asai Y، و همکاران. انتقال مکانیکی در سلولهای موی گوش داخلی موش به ژنهای کانالمانند غشایی نیاز دارد.J Clin Invest. 2011.121:4796 809. doi: 10.1172/JCI60405.�[PMC رایگان مقاله]�[گروه]�[صلیب نماینده]
92. تلیلی ع، ربه آیبی، عیفا-همانی م، ذویب ح، معلا ج، تلیلی-چوچنه ج، و همکاران. TMC1 اما نه TMC2 مسئول اختلالات شنوایی غیر سندرمی اتوزومال مغلوب در خانواده های تونسی است.Audiol Neurootol.�2008.13: 213 8. doi: 10.1159 / 000115430. [گروه]�[صلیب نماینده]
93. Manji SS، Miller KA، Williams LH، Dahl HH. شناسایی سه سویه موش کم شنوایی جدید با جهش در ژن Tmc1.جی پاتول هستم.2012.180: 1560 9. doi: 10.1016/j.ajpath.2011.12.034.[گروه]�[صلیب نماینده]
94. Wetzel C، Hu J، Riethmacher D، Benckendorff A، Harder L، Eilers A، و همکاران. یک پروتئین حوزه استوماتین ضروری برای حس لامسه در موش.طبیعت . 2007.445: 206. doi: 9/nature10.1038.�[گروه][صلیب نماینده]
95. Martinez-Salgado C، Benckendorff AG، Chiang LY، Wang R، Milenkovic N، Wetzel C، و همکاران. استوماتین و انتقال مکانیکی نورون حسی.�J Neurophysiol. 2007.98: 3802~8. doi: 10.1152/jn.00860.2007.[گروه]�[صلیب نماینده]
96. Huang M، Gu G، Ferguson EL، Chalfie M. یک پروتئین شبه استوماتین لازم برای ایجاد حس مکانیکی در C. elegans.طبیعت . 1995.378:292�5. doi: 10.1038/378292a0.�[گروه]�[صلیب نماینده]
97. هو جی، چیانگ لی، کوچ ام، لوین GR. شواهدی برای اتصال پروتئینی درگیر در لمس جسمی.EMBO J. 2010.29: 855. doi: 67/emboj.10.1038.�[PMC رایگان مقاله]�[گروه]�[صلیب نماینده]
98. Chiang LY، Poole K، Oliveira BE، Duarte N، Sierra YA، Bruckner-Tuderman L، و همکاران. لامینین-332 انتقال مکانیکی و انشعاب مخروط رشد را در نورون های حسی هماهنگ می کند.Nat Neurosci. 2011.14: 993. doi: 1000/nn.10.1038.�[گروه]�[صلیب نماینده]
99. Lesage F، Guillemare E، Fink M، Duprat F، Lazdunski M، Romey G، و همکاران. TWIK-1، یک انسان در همه جا حاضر با تصحیح کانال K+ ضعیف به سمت داخل با ساختاری جدید.EMBO J. 1996.15: 1004 11.[PMC رایگان مقاله]�[گروه]
100. Lesage F. فارماکولوژی کانال های پتاسیم پس زمینه عصبی.�نوروفارماکولوژی.�2003.44:1�7. doi: 10.1016/S0028-3908(02)00339-8.�[Review]�[گروه]�[صلیب نماینده]
101. Medhurst AD، Rennie G، Chapman CG، Meadows H، Duckworth MD، Kelsell RE، و همکاران. تجزیه و تحلیل توزیع کانالهای پتاسیم دو حفرهای انسانی در بافتهای سیستم عصبی مرکزی و محیطی.Brain Res Mol Brain Res.�2001.86:101�14. doi: 10.1016/S0169-328X(00)00263-1.�[گروه][صلیب نماینده]
102. Maingret F، Patel AJ، Lesage F، Lazdunski M، Honor E. تحریک مکانیکی یا اسیدی، دو حالت تعاملی فعال سازی کانال پتاسیم TREK-1.J Biol Chem. 1999.274: 26691. doi: 6/jbc.10.1074.[گروه]�[صلیب نماینده]
104. Alloui A، Zimmermann K، Mamet J، Duprat F، No'l J، Chemin J، و همکاران. TREK-1، یک کانال K+ که در درک درد چندوجهی نقش دارد.EMBO J. 2006.25: 2368 76. doi: 10.1038/sj.emboj.7601116.[PMC رایگان مقاله]�[گروه]�[صلیب نماینده]
105. نول جی، زیمرمن کی، بوسرولز جی، دیوال ای، آلویی آ، دیوشوت اس، و همکاران. کانال های K+ فعال شده مکانیکی TRAAK و TREK-1 هم ادراک گرم و هم سرد را کنترل می کنند.EMBO J. 2009.28: 1308. doi: 18/emboj.10.1038.�[PMC رایگان مقاله]�[گروه]�[صلیب نماینده]
106. Dobler T، Springauf A، Tovornik S، Weber M، Schmitt A، Sedlmeier R، و همکاران. کانالهای K+ دو حفرهای TRESK جزء مهمی از جریانهای پتاسیم پسزمینه در نورونهای گانگلیون ریشه پشتی موش را تشکیل میدهند.J Physiol. 2007.585: 867 79. doi: 10.1113/jphysiol.2007.145649.[PMC رایگان مقاله]�[گروه]�[صلیب نماینده]
107. Bautista DM، Sigal YM، Milstein AD، Garrison JL، Zorn JA، Tsuruda PR، و همکاران. عوامل تند فلفل Szechuan با مهار کانالهای پتاسیم دو منافذ، نورونهای حسی را تحریک میکنند.Nat Neurosci. 2008.11: 772. doi: 9/nn.10.1038.�[PMC رایگان مقاله]�[گروه]�[صلیب نماینده]
109. Heidenreich M، Lechner SG، Vardanyan V، Wetzel C، Cremers CW، De Leenheer EM، و همکاران. کانال KCNQ4 K(+) گیرنده های مکانیکی را برای حس لمس طبیعی در موش و انسان تنظیم می کند.Nat Neurosci. 2012.15: 138. doi: 45/nn.10.1038.�[گروه]�[صلیب نماینده]
110. فرنزل اچ، بوهلندر جی، پینسکر کی، وولبن بی، تانک جی، لچنر اس جی، و همکاران. یک مبنای ژنتیکی برای صفات حسی مکانیکی در انسانPLoS Biol.�2012.10: e1001318. doi: 10.1371 / journal.pbio.1001318.[PMC رایگان مقاله]�[گروه]�[صلیب نماینده]
درد مکانیکی: با توجه به CDC، بیش از 50٪ از بزرگسالان ایالات متحده (125 میلیون) دارای اسکلتی عضلانی بودند. اختلال درد در سال 2012.
بیش از 40 درصد از بزرگسالان مبتلا به اختلال درد اسکلتی عضلانی در سال 2012 به هر دلیلی از رویکرد سلامت مکمل استفاده کردند. استفاده از رویکردهای بهداشتی مکمل به هر دلیلی در بین افراد مبتلا به گردن درد یا مشکلات بیش از دو برابر استفاده در افراد بدون این مشکلات بود.
در میان بزرگسالان مبتلا به اختلال درد اسکلتی عضلانی، استفاده از هر رویکرد بهداشتی مکمل در میان افراد مبتلا به گردن درد یا مشکلات (50.6 درصد) و پس از آن افراد با سایر مشکلات اسکلتی عضلانی (46.2 درصد) بالاترین میزان بود.
استفاده از رویکردهای بهداشتی مکمل به هر دلیلی در میان افراد مبتلا به گردن درد یا مشکلات بیش از دو برابر استفاده در افراد بدون این مشکلات بود.
گیرنده های مکانیکی گیرنده های حسی هستند که به فشار یا اعوجاج مکانیکی پاسخ می دهند.
اینها شامل گیرنده های پوستی برای لمس، گیرنده هایی که طول و تنش ماهیچه ها را کنترل می کنند، گیرنده های شنوایی و دهلیزی و غیره هستند.
تئوری درد کنترل دروازه
ورودی غیر دردناک دروازه ها را به ورودی دردناک می بندد.
این مانع از انتقال درد به سطوح بالاتر قشر مغز می شود
آوران های با قطر کوچک (درد) مانع از مهار درد می شوند
آوران های با قطر زیاد (ارتعاش) تمایل به تحریک مهار درد دارند.
این نظریه ادعا میکند که فیبرهای غیردردی میتوانند با سیگنالهای فیبرهای درد تداخل داشته باشند، بنابراین، درد را مهار میکنند.
قطر بزرگ A الیاف غیردردی هستند (محرک های درد را منتقل نمی کنند) و اثرات شلیک توسط A را مهار می کنند؟ و الیاف C
مسیر لمنیسکال میانی ستون پشتی
استفاده از گیرنده های مکانیکی محیطی برای تغییر ادراک درد
چگونه می توانیم کمک کنید؟
رفلکس عقب نشینی
یک نورون آوران برانگیخته، نورونهای بینالمللی تحریکی را تحریک میکند که به نوبه خود نورونهای حرکتی وابران تامینکننده عضله دوسر بازو، ماهیچهای در بازو که مفصل آرنج را خم میکند، تحریک میکند. انقباض عضله دوسر بازو، دست را از اجاق داغ دور می کند.
نورون آوران همچنین نورون های بازدارنده ای را تحریک می کند که به نوبه خود نورون های وابران تامین کننده عضلات سه سر را مهار می کنند تا از انقباض آن جلوگیری کنند. این نوع اتصال عصبی که شامل تحریک تامین عصب به یک عضله و مهار همزمان اعصاب به عضله متضاد آن است، به عنوان مهار متقابل شناخته می شود.
نورون آوران هنوز هم سایر نورون های داخلی را تحریک می کند که سیگنال را از طریق یک مسیر صعودی از طناب نخاعی به مغز می برند. تنها زمانی که تکانه به ناحیه حسی قشر می رسد، فرد از درد، محل آن و نوع محرک آگاه می شود. علاوه بر این، هنگامی که تکانه به مغز می رسد، اطلاعات می تواند به عنوان حافظه ذخیره شود و فرد می تواند درباره آنچه اتفاق افتاده فکر کند.
درمان مبتنی بر گیرنده
تنظیمات
فعالسازی گیرندههای مکانیکی مفصلی از طریق تنظیمات کایروپراکتیک میتواند درک مغز از الیاف با قطر کوچکتر را تعدیل کرده و « تحت الشعاع» قرار دهد.
تکرار فعال شدن گیرندههای مکانیکی مفصلی میتواند شکل پذیری مثبتی در مسیرهای آوران ایجاد کند.
انعطاف پذیری مثبت می تواند درد را خاموش کند
ارتعاش
تحریک ارتعاشی در فرکانس های خاص می تواند درک درد را تغییر دهد
تکرار فعالسازی دیسکهای مرکل و سلولهای مایسنر میتواند انعطافپذیری مثبتی در مسیرهای آوران ایجاد کند.
باز هم، انعطاف پذیری مثبت می تواند درد را خاموش کند
ارتعاش
این نوع دستگاه ارتعاشات سینوسی را اعمال می کند و دامنه قابل انتخاب پیوسته 0-5.2 میلی متر بسته به موقعیت پا و فرکانس قابل انتخاب 5-30 هرتز را ارائه می دهد.
به نظر می رسد آموزش WBV یک مداخله موثر، ایمن و مناسب برای کارکنان نشسته با کمردرد مزمن باشد.
تحریک ارتعاشی-لمسی هموتوپیک منجر به کاهش 40 درصدی درد گرمایی در همه گروهها شد. به نظر نمی رسید حواس پرتی بر رتبه بندی درد تجربی تأثیر بگذارد
"تحریک لامسه ارتعاشی نه تنها در NC بلکه در بیماران مبتلا به درد مزمن اسکلتی عضلانی، از جمله FM، به طور موثر مکانیسم های ضد درد را به کار می گیرد."
لمس نور
درمجموع، 44 داوطلب سالم، درد گرمایی و CT بهینه (مسواک زدن آهسته) و CT غیربهینه (مسواک زدن سریع یا لرزش) را تجربه کردند. سه الگوی تجربی مختلف مورد استفاده قرار گرفت: کاربرد همزمان درد گرما و تحریک لمسی (مسواک زدن آهسته یا ارتعاش). مسواک زدن آهسته، برای مدت زمان و فواصل متغیر، قبل از درد گرما اعمال می شود. مسواک زدن آهسته در مقابل سریع مسواک زدن قبل از درد گرما.
در انسان، نواحی اصلی مغز که اطلاعات C-LTMR را دریافت میکنند، متعلق به سیستم حسی تنی هستند و بر روی شبکههای مغزی مانند قشر منزوی خلفی طرف مقابل یا قشر پیش پیشانی میانی تأثیر میگذارند. شدت لمس هدفمند CT در قشر حسی سوماتوی اولیه و ثانویه (S1 طرف مقابل، S2 دو طرفه) کدگذاری میشود، در حالی که لذت در قشر کمربندی قدامی کدگذاری میشود. C-LTMR ها همچنین نواحی درگیر در پردازش پاداش (پوتامن و قشر اوربیتوفرونتال) و در پردازش محرک های اجتماعی (شیار گیجگاهی فوقانی خلفی) را فعال می کنند.
هر چیز محیطی یک پیامد مرکزی دارد
مطالعه موردی
مرد 47 ساله در اکتبر 2017 دچار CVA چپ شد.
بعد از تصادف سمت راست بدنش را تکان نداده است.
به کلینیک ما ارائه شد زیرا میخواهد «به آن برگردد».
نکات برجسته معاینه فیزیکی
اختلال تکلم
تغییر ادراک درد
مشکل با ریاضیات ساده
شل در RUE و RLE
نکات برجسته معاینه فیزیکی
بیمار هیچ حرکتی نداشت تا اینکه ما شروع به آزمایش حس و رفلکس کردیم:
آلوداینیا:به حساسیت درد مرکزی (افزایش پاسخ نورون ها) به دنبال تحریک معمولی غیردردناک و اغلب تکراری اشاره دارد.
آلوداینیا میتواند منجر به تحریک پاسخ درد از محرکها شود که معمولاً باعث تحریک درد نمیشود.
دما یا محرکهای فیزیکی میتوانند آلوداینیا را تحریک کنند، که ممکن است مانند یک احساس سوزش باشد و اغلب پس از آسیب به یک محل رخ میدهد.
Alldynia با Hyperalgesia، یک واکنش افراطی و اغراق آمیز به یک محرک، که معمولاً دردناک است، متفاوت است.
چرا آسیب یا آسیب ناشی از تروما موضعی سبب درد مزمن و غیر قابل تحمل در بیماران خاص می شود؟ چه اتفاقی برای ترجمه آسیب های موضعی با درد حاد به یک وضعیت مزمن رنج می افتد؟ چرا برخی از درد به داروهای ضد التهابی و / یا داروها پاسخ می دهند، در حالی که دیگر انواع درد نیاز به مواد مخدر دارند؟
درد یک فرایند پیچیده شامل سیستم عصبی محیطی (PNS) و سیستم عصبی مرکزی (CNS) است. آسيب بافت باعث پيدايش PNS مي شود که سيگنال ها را از طريق نخاع به مغز منتقل مي کند که در آن درک درد اتفاق مي افتد. با این حال، چه چیزی موجب شد تا درد شدید درد به پدیده ی بی وقفه تبدیل شود؟ آیا می توان برای جلوگیری از آن کاری انجام داد؟ شواهد نشان می دهد که درد مزمن نتایج حاصل از ترکیبی از مکانیزم، مانند "خاطرات" عصبی از درد قبل.
ناسازگاری: ساده ترین راه
درد حاد و یا درد ناخوشایندی به عنوان تجربه معمولی ناراحتی مشخص می شود که در پاسخ به آسیب های شدید اولیه یا آسیب دیده رخ می دهد. این محافظ است و ما را هشدار می دهد تا از ابتلا به این توهین دور شویم و از تروما مراقبت کنیم. مکانیسم هایی که باعث درد ناخوشایندی می شوند، شامل انتقال، که تحرک آسیب دیده خارجی را به فعالیت های الکتریکی در اعصاب مرکزی متناوب غیر تخصصی متصل می کند. اعصاب عصبی سپس اطلاعات حسی را از PNS به CNS منتقل می کنند.
در CNS، داده های درد توسط نورون های حسی اولیه به سلول های پروژکتور مرکزی منتقل می شوند. پس از انتقال اطلاعات به تمامی مناطقی از مغز که مسئول ادراک ما هستند، تجربه حسی واقعی اتفاق می افتد. درد ناخوشی یک واکنش نسبتا ساده به یک محرک بسیار ساده و حاد است. اما مکانیک مسئول درد درد ناخوشایند نمی تواند پدیده هایی را پیدا کند، مانند درد که با وجود حذف یا بهبود تحریک، همچون در مورد درد اندام فانتوم، همچنان ادامه دارد.
درد و پاسخ التهابی
در شرایطی که آسیب شدیدتر، مانند زخم های جراحی، آسیب بافت ممکن است واکنش التهابی را تحریک کند. با این حال، شرایط دیگر، به ویژه آرتریت، همچنین می تواند با موارد مداوم التهاب مرتبط با علائم شدید درد مشخص شود. مکانیسم های این نوع درد که مربوط به آسیب بافتی و پاسخ التهابی است متفاوت از درد زودهنگام هشدار دهنده است.
در صورت بروز نقص یا محل آسیب یا آسیب دیگر، یک حادثه از حوادث غیر قابل انقباض در سیستم عصبی رخ می دهد. این پدیده "بادی" از بدن آغاز می شود در پوست، جایی که آن را در امتداد اعصاب محیطی تقویت می کند، و در پاسخ حساسیت بالا در امتداد نخاع (شاخ پشتی) و مغز به دست می آید. سپس سلولهای التهابی مناطق آسیب بافتی را احاطه کرده و همچنین سیتوکین ها و کیموکان ها را تولید می کنند که مواد اصلی آن در بین فرایندهای بهبود و بازسازی بافت ها هستند. اما، این عوامل همچنین ممکن است باعث تحریک کننده و تنظیم خواص نورون های حسی اولیه اطراف منطقه تروما شوند.
بنابراین، عوامل اصلی که موجب درد التهابی می شوند شامل آسیب به ناخوشی های آستانه بالا آستیگماتیسم شناخته شده به عنوان حساسیت محیطی، تغییرات و تغییرات نورون در سیستم عصبی و تقویت تحریک پذیری نورون درون CNS می باشد. این نشان دهنده حساسیت مرکزی است و برای حساسیت بالا پاسخگو است، در صورتی که در مناطقی که مجاور آسیب های واقعی هستند، درد زخمی خواهد شد. این بافت ها همچنین می توانند به تحریک واکنش نشان دهند که به طور معمول درد، مانند لمس، پوشیدن لباس، فشار نور و حتی مسواک زدن موهای خود را ایجاد نمی کند، مثل اینکه واقعا دردناک است و به عنوان آلدینیا شناخته می شود.
درد نوروپاتیک ناشی از آسیب یا آسیب به سیستم عصبی، مانند سندرم تونل کارپ، نورالژی پس از فروپتیک و نوروپاتی دیابتی است. اگر چه بعضی از مکانیسم هایی که به نظر می رسد باعث درد نوروپاتی می شوند، با کسانی که علت درد التهابی را درگیر می کنند، بسیاری از آنها متفاوت هستند و به همین دلیل نیازمند رویکرد متفاوت نسبت به مدیریت آنها هستند.
فرایند حساسیت محیطی و مرکزی، حداقل از لحاظ تئوری و تجربی، در طول انتقال دهنده عصبی، گلوتامات، که گمان می رود آزاد شدن زمانی که گیرنده N-methyl-D-aspartate (NMDA) فعال می شود، حفظ می شود.
سیستم عصبی ساخته شده از هر دو انتقال دهنده های عصبی مهار کننده یا تحریک کننده است. اکثر آنچه که سیستم عصبی ما را قادر به پاسخ مناسب به آسیب یا آسیب می کند، تنظیم یا مهار فرآیندهای مختلف است. بیش از حد تحریک سیستم عصبی در بسیاری از اختلالات متفاوت دیده می شود. به عنوان مثال، بیش از حد فعال شدن گیرنده NMDA نیز می تواند به اختلالات عاطفی، اختلالات سمپاتیک و حتی تحمل به مواد مخدر مرتبط باشد.
حتی درد ناخوشایندی عادی، تا حدودی، گیرنده NMDA را فعال می کند و بر این باور است که منجر به انتشار گلوتامات می شود. با این حال، در درد نوروپاتیک، حساسیت به گیرنده NMDA کلید است.
با سایر انواع درد مزمن، مانند فیبرومیالژیا و سردرد تنشي، برخی از مکانیزم های فعال در درد التهابی و نوروپاتیک همچنین ممکن است اختلالات مشابهی در سیستم درد ایجاد کنند، از جمله حساسیت مرکزی، تحریک پذیری بالاتر مسیرهای سموتوسنسوری و کاهش مکانیزم مهار کننده سیستم عصبی مرکزی.
حساسیت محیطی
Cyclo-oxygenase (COX) نیز نقش مهمی در حساسیت محیطی و مرکزی دارد. COX-2 یکی از آنزیم هایی است که در طول پروسه التهابی ایجاد می شود. COX-2 اسید اورهیدونیک را به پروستاگلاندین ها تبدیل می کند که باعث افزایش حساسیت ترمینال های محیطی ناخوشایند می شود. عملا، التهاب محیطی باعث می شود COX-2 از CNS تولید شود. سیگنال های نئوسیتپتورهای محیطی بخشی از مسئولیت این تنظیم است، اما به نظر می رسد یک عنصر humoral برای انتقال سیگنال های درد در سراسر مانع خون مغزی.
به عنوان مثال، در مدل های تجربی، COX-2 از CNS تولید می شود، حتی اگر حیوانات قبل از تحریک التهابی محیطی، بلوک عصبی حسی را دریافت می کنند. COX-2 که بر روی نورون های شاخ پشتی نخاعی بیان می شود، پروستاگلاندین ها را که بر پایانه های مرکزی یا پایانه های پیشیناپتیک فیبرهای حسی غوطهور عمل می کنند، آزاد می کند. علاوه بر این، آنها به طور Postynaptically در نورون شاخ پشتی به طور مستقیم depolarization تولید. و در نهایت، آنها فعالیت گلیکول گلیسین را مهار می کنند، و این یک فرستنده مهار کننده است. بنابراین، پروستاگلاندین ها باعث افزایش تحریک پذیری نورون های مرکزی می شوند.
پلاستیسیته مغز و حساسیت مرکزی
حساسیت مرکزی تغییراتی را که در مغز در واکنش به تحریک مکرر عصب اتفاق می افتد توصیف می کند. بعد از محرک های مکرر، مقدار هورمون ها و سیگنال های الکتریکی مغز تغییر می کند به این ترتیب نورون ها "حافظه" را برای واکنش به این نشانه ها ایجاد می کنند. تحریک ثابت باعث ایجاد حافظه قوی تر مغز می شود، به طوری که مغز سریع و مؤثرتر در صورت تحریک یکسان در آینده پاسخ خواهد داد. تغییرات در نتیجه در سیم کشی و واکنش مغزی به عنوان پلاستیک عصبی نامیده می شود که توانایی مغز را به راحتی تغییر می دهد یا حساسیت مرکزی را توصیف می کند. بنابراین مغز با محرک های قبلی یا مکرر فعال شده و حساسیت می یابد تا بیشتر تحریک پذیر شود.
نوسانات حساسیت مرکزی پس از برخورد مکرر با درد رخ می دهد. تحقیقات در حیوانات نشان می دهد که قرار گرفتن در معرض مکرر به تحریک دردناک، آستانه درد را در حیوان تغییر می دهد و منجر به پاسخ قوی تر درد می شود. محققان معتقدند که این تغییرات می تواند درد مداوم را که حتی پس از عمل جراحی پشتی نیز رخ می دهد، توضیح دهد. اگر چه یک دیسک فتق ممکن است از یک عصب خاردار برداشته شود، درد ممکن است به عنوان حافظه فشرده سازی عصب ادامه یابد. نوزادان تحت درمان با ختنه بدون بیهوشی عمیق تر به تحریک دردناک آینده، مانند تزریق روتین، واکسیناسیون و سایر فرآیندهای دردناک واکنش نشان می دهند. این کودکان نه تنها یک واکنش همودینامیک بالاتر دارند، که به عنوان تاکی کاردی و تاکیپنای شناخته می شوند، بلکه همچنین افزایش گریه نیز افزایش می یابد.
این حافظه عصبی درد شدیدا مورد مطالعه قرار گرفت. Woolf در گزارشی در مورد مطالعات قبلی تحقیقاتی خود خاطرنشان کرد که تحریک پذیری بهبود یافته رفلکس پس از آسیب یا آسیب بافتی محیطی بر روی ادامه سیگنال های ورودی محیطی تکیه نمی کند؛ در عوض، ساعت ها پس از یک آسیب محیطی، میدان مغناطیسی نورون سوراخ نخاعی عقب ادامه یافت. محققان همچنین اهمیت گیرنده NMDA نخاعی را برای القاء و حفظ حساسیت مرکزی ثبت کرده اند.
اهمیت مدیریت درد
هنگامی که حساسیت مرکزی ایجاد می شود، دوزهای بزرگ ضد درد اغلب لازم است تا آن را سرکوب کنند. مقاربت پیشگیرانه یا درمان قبل از پیشرفت درد، ممکن است اثرات همه این تحریکات بر روی CNS را کاهش دهد. وولف نشان داد که دوز مورفین مورد نیاز برای متوقف نمودن غلظت بالای مغز، که قبل از تحریک الکتریکی کوتاه مدت در موش صحرایی داده می شد، یک دهم دوز لازم برای از بین بردن فعالیت پس از رشد بود. این به عمل بالینی ترجمه می شود.
در یک کارآزمایی بالینی بیماران 60 که تحت هیسترکتومی شکمی قرار گرفتند، افرادی که در زمان تزریق بیهوشی به صورت داخل وریدی 10 میلی گرم مورفین دریافت کردند، برای کنترل درد پس از عمل، مورفین به میزان قابل توجهی کمتر بود. علاوه بر این، حساسیت درد در اطراف زخم، که به عنوان hyperalgesia ثانویه نامیده می شود، نیز در گروه پیشگیری شده مورفین کاهش می یابد. مقاربت پیشگیرانه با موفقیت قابل مقایسه در مجموعه ای از تنظیمات جراحی، از جمله عمل جراحی و عمل جراحی پس از عمل جراحی استفاده شده است.
دوز منفرد 40 یا 60 mg / kg استامینوفن رکتال دارای اثر مؤثری مورفین در جراحی روزانه در کودکان است که در صورت استفاده از بیهوشی تزریق می شود. علاوه بر این، کودکان مبتلا به کمبودهای شدید با استامینوفن، تهوع و استفراغ پس از عمل به میزان قابل توجهی کمتر می شوند.
آنتاگونیست های گیرنده NMDA هنگام بیهوشی قبل از عمل، درد پس از عمل را تحمل می کنند. گزارش های مختلف در ادبیات وجود دارد که حاکی از استفاده از کتامین و دکسترومتورفان در دوره پیش از عمل است. در بیماران تحت بازسازی رباط صلیبی قدامی، میزان مصرف مواد مخدر از نظر درد بیمار تحت کنترل 24 ساعت به طور معنی داری در گروه قبل از عمل جراحی دکسترومتورفان در مقایسه با گروه پلاسبو کمتر بود.
در مطالعات دوسوکور، کنترل شده با پلاسبو، گاباپنتین به عنوان یک داروی ضد درد برای بیمارانی که تحت عمل جراحی ماسستکتومی و هیسترکتومی قرار گرفته بودند، نشان داده شد. گاباپنتین خوراکی قبل از عمل، نمرات درد و مصرف داروهای ضد درد را بدون شکاف در عوارض جانبی کاهش می دهد، در مقایسه با دارونما.
تجویز قبل از عمل از داروهای ضد التهاب غیر استروئیدی (NSAIDs) کاهش قابل توجهی در مصرف مواد مخدر پس از عمل نشان داده است. COX-2 ها به دلیل کمبود نسبی عوارض پلاکتی و پروفایل ایمنی معده در مقایسه با NSAID های معمول ترجیح داده می شوند. سلکوکسیب، روتوکوکسیب، دارونکوکسیب و پارکوکسیب، خارج از ایالات متحده، پیش از عمل، مصرف مواد مخدر بعد از عمل را با بیش از 40 درصد کاهش داد، و بسیاری از بیماران از کمتر از نصف مواد مخدر در مقایسه با دارونما استفاده می کردند.
به نظر می رسد مسدود کردن هدایت عصبی در دوره قبل از عمل از ایجاد حساسیت مرکزی جلوگیری می کند. سندرم اندام فانتوم (PLS) به یک پدیده باد کردن ستون فقرات نسبت داده شده است. بیماران دچار قطع عضو
اغلب سوزش یا درد سوزن در بخش بدن از بین رفته است. یکی از علل احتمالی این است که فیبرهای عصبی در ناحیه تنه تحریک می شوند و مغز سیگنال ها را به عنوان مقدمه ای در بخش انتصاب شده تفسیر می کند. دیگر بازسازی در داخل مناطق قشر است به طوری که منطقه برای دست می گوید در حال حاضر به سیگنال های دیگر از بدن پاسخ می دهد، اما هنوز آنها را تفسیر به عنوان آمدن دست دست انداز است.
با این حال، برای بیماران تحت قطع عضو اندام تحتانی تحت بیهوشی اپیدورال، هیچ یک از بیماران 11 که بلوک اپیدورال کمری را با بوپیواکائین و مورفین برای 72 ساعت قبل از عمل دریافت کردند، PLS را توسعه دادند. برای افرادی که بیهوشی عمومی بدون بلوک اپیدورال کمری داشتند، 5 از بیماران 14 دارای PLS در هفته 6 بود و 3 همچنان در سال 1 PLS را تجربه می کرد.
وولف و چانگ اشاره کرده اند که درمان کامل قبل از عمل جراحی، عمل جراحی و بعد از عمل شامل "NSAID ها برای کاهش فعال شدن / متمرکز کردن نقص ایمنی، بیهوشی موضعی برای جلوگیری از جریان حسی و داروهای مرکزی عمل مانند مواد مخدر است." کاهش درد درازمدت با تکنیک های پیشگیرانه افزایش می یابد رضایت، تنگی تنگ، مصرف مواد مخدر، همراه با یبوست کاهش یافته، آرامبخش، حالت تهوع، و احتباس ادراری، و حتی ممکن است باعث ایجاد درد مزمن شود. متخصصین بیهوشی و جراحان باید این تکنیک ها را در شیوه های روزمره خود در نظر بگیرند.
هنگامی که درد ناشی از آسیب یا آسیب در نتیجه عمل جراحی اتفاق می افتد، نخاع می تواند به حالت غربالگری منتهی شود که در آن واکنش های درد بیش از حد رخ می دهد که ممکن است برای روزها، هفته ها یا حتی سال ها ادامه یابد.
چرا آسیب موضعی ناشی از تروما باعث درد مزمن و غیر قابل تحمل در برخی از بیماران می شود؟ آسيب بافت منجر به صورت فلکی تغييرات تحريکی ستون فقرات می شود، از جمله شلیک خود به خودی، دامنه و طول پاسخ بیشتر، کاهش آستانه، افزایش ترشح برای تحریک مکرر و افزایش میدان های پذیرش. پایداری این تغییرات که به طور کلی حساسیت مرکزی نامیده می شود، به نظر می رسد که برای افزایش طولانی مدت حساسیت درد است که درد مزمن را تعیین می کند. داروهای متعدد و یا داروها و همچنین بلوک عصبی بیهوشی محلی ممکن است موجب ایجاد سرگیجه سیستم عصبی مرکزی (CNS) را محدود کند، که نشان دهنده کاهش درد و کاهش مصرف مواد مخدر در مدل های ضد درد پیشگیرانه است.
دکتر الکس جیمنز Insight
مراقبت از کایروپراکتیک یک گزینه درمان جایگزین است که از تنظیمات اسپینال و دستکاری دستی برای ایمن و موثر بازگرداندن و همچنین حفظ هماهنگی مناسب ستون فقرات استفاده می کند. تحقیقات تحقیقاتی نشان داده اند که عدم تعادل ستون فقرات یا زیرولفکساسیون می تواند منجر به درد مزمن شود. مراقبت های کایروپراکتیک معمولا برای مدیریت درد استفاده می شود، حتی اگر نشانه ها به آسیب و / یا وضعیت سیستم عضلانی اسکلتی و عصبی مرتبط نباشد. با دقت تنظیم مجدد ستون فقرات، یک کیهان شناسی می تواند به کاهش استرس و فشار از سازه های اطراف اجزای اصلی پایه بدن کمک کند، در نهایت تسکین درد را کاهش می دهد.
عملکرد سیستم عصبی در سیستم عصبی و درد
هنگامی که به کاهش استفاده از داروها و / یا داروها، از جمله مواد افیونی کمک می کند، به منظور جلوگیری از عوارض جانبی مانند مسائل مربوط به سلامت دستگاه گوارش، عملکرد مناسب سیستم عصبی قلبی ممکن است در بازی باشد.
سیستم عصبی روده (ENS) یا سیستم عصبی ذاتی یکی از شاخه های کلیدی سیستم عصبی خودمختار (ANS) است و از یک سیستم شبکه مانند اعصاب تشکیل شده است که نقش دستگاه گوارش را تعدیل می کند. این می تواند مستقل از سیستم عصبی سمپاتیک و پاراسمپاتیک عمل کند، حتی اگر ممکن است تحت تأثیر آنها قرار گیرد. ENS را می توان مغز دوم نیز نامید. از سلول های تاج عصبی مشتق شده است.
سیستم عصبی عصبی در انسان از برخی از نورونهای 500 میلیون ساخته شده است، از جمله انواع متعددی از سلول های Dogiel، تقریبا یک دوسوم از مقدار نورون در مغز. سیستم عصبی عصبی به داخل پوشش سیستم دستگاه گوارش وارد می شود و از مری شروع می شود و تا انتهای آن گسترش می یابد. سلول های Dogiel، همچنین به عنوان سلول های Dogiel شناخته می شود، به نوعی از بافت های فوقانی چند قطبی داخل گانگلیس سمپاتیک پیش پرده می گویند.
ENS قادر به انجام عملکردهای مستقل مانند هماهنگی رفلکس ها است. حتی اگر عصب قابل توجهی در سیستم عصبی خودمختار دریافت می کند، مستقل از مغز و نخاع عمل می کند و می تواند عمل کند. سیستم عصبی روده به دلایل متعددی به عنوان "مغز دوم" توصیف شده است. سیستم عصبی روده ممکن است به طور مستقل عمل کند. به طور معمول با سیستم عصبی مرکزی (CNS) از طریق پاراسمپاتیک، یا از طریق عصب واگ، و سمپاتیک، یعنی از طریق گانگلیون های پیش مهره ای، سیستم عصبی ارتباط برقرار می کند. با این حال، مطالعات مهرهداران نشان میدهد که وقتی عصب واگ قطع میشود، سیستم عصبی روده به کار خود ادامه میدهد.
در مهره داران، سیستم عصبی عصبی شامل نورون های مؤثر، نورون های عصبی و اینترنورون ها می شود که این همه سیستم عصبی روده را قادر به انجام رفلکس می کند و در صورت عدم ورودی CNS به عنوان یک مرکز تلفیقی عمل می کند. نورونهای حسی به شرایط مکانیکی و شیمیایی گزارش می دهند. سیستم عصبی عصبی توانایی تغییر واکنش خود را بر اساس عوامل مانند ترکیب مواد مغذی و فله ای دارد. علاوه بر این، ENS شامل سلول های پشتیبانی می شود که بسیار شبیه آستروگلیا مغز و مانع نفوذی در اطراف مویرگ های اطراف انقباضات است که مانع انسداد خون مغزی است.
سیستم عصبی روده (ENS) نقش اساسی در فرآیندهای التهابی و درد دارد. داروها و/یا داروهایی که با ENS تداخل دارند اخیراً به دلیل توانایی آنها در تنظیم جنبه های متعدد فیزیولوژی روده و پاتوفیزیولوژی مورد توجه قابل توجهی قرار گرفته اند. به طور خاص، آزمایشها روی حیوانات نشان دادهاند که گیرندههای فعال شده با پروتئیناز (PARs) ممکن است برای التهاب عصبی در روده ضروری باشند. علاوه بر این، به نظر میرسد آگونیستهای PAR2 باعث ایجاد حساسیت و حالتهای پردردی روده میشوند که نقش این گیرنده را در درک درد احشایی نشان میدهد.
علاوه بر این، PAR ها، همراه با پروتئینازهایی که آنها را فعال می کنند، اهداف جدید هیجان انگیز را برای مداخله درمانی در ENS نشان می دهند. محدوده اطلاعات ما محدود به کیهان شناسی و همچنین آسیب های ناشی از ستون فقرات و شرایط است. برای بحث در مورد موضوع، لطفا از دکتر جیمنز بپرسید یا با ما تماس بگیرید 915-850-0900 .
دکتر الکس جیمنز سرپرستی می کند
موارد اضافی: سیاتیک
سیاتیک از نظر پزشکی به عنوان مجموعه ای از علائم شناخته شده است، نه یک آسیب و / یا یک وضعیت واحد. علائم درد عصب سیاتیک یا سیاتیک می تواند با شدت و فرکانس متفاوت باشد، اما اغلب به عنوان ناگهانی، چاقو یا درد الکتریکی توصیف می شود که از پایین کمر عقب پایین باسن، باسن، ران و پاها را به پای سایر علائم سیاتیک ممکن است شامل احساس سوزن شدن یا سوزش، بی حسی و ضعف در طول طول عصب سیاتیک باشد. سیاتیک اغلب افراد سالهای 30 و 50 را تحت تاثیر قرار می دهد. این ممکن است اغلب به علت انحطاط ستون فقرات به علت سن ایجاد شود، اما فشرده سازی و تحریک عصب سیاتیک ناشی از انفجار یا دیسک حنجره، در میان سایر مسائل مربوط به سلامت ستون فقرات، همچنین ممکن است درد عصب سیاتیک ایجاد کند.
IFM's Find A Practitioner بزرگترین شبکه ارجاع در پزشکی کاربردی است که برای کمک به بیماران در یافتن پزشکان طب عملکردی در هر نقطه از جهان ایجاد شده است. با توجه به تحصیلات گسترده در پزشکی کاربردی ، پزشکان مجاز IFM اولین بار در نتایج جستجو ذکر شده اند

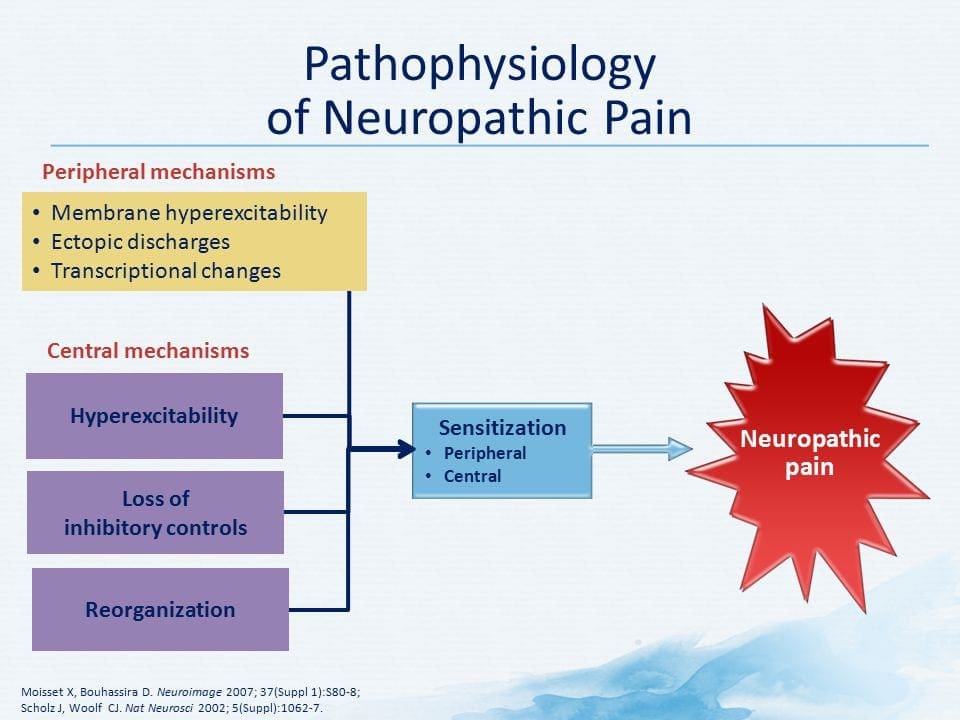
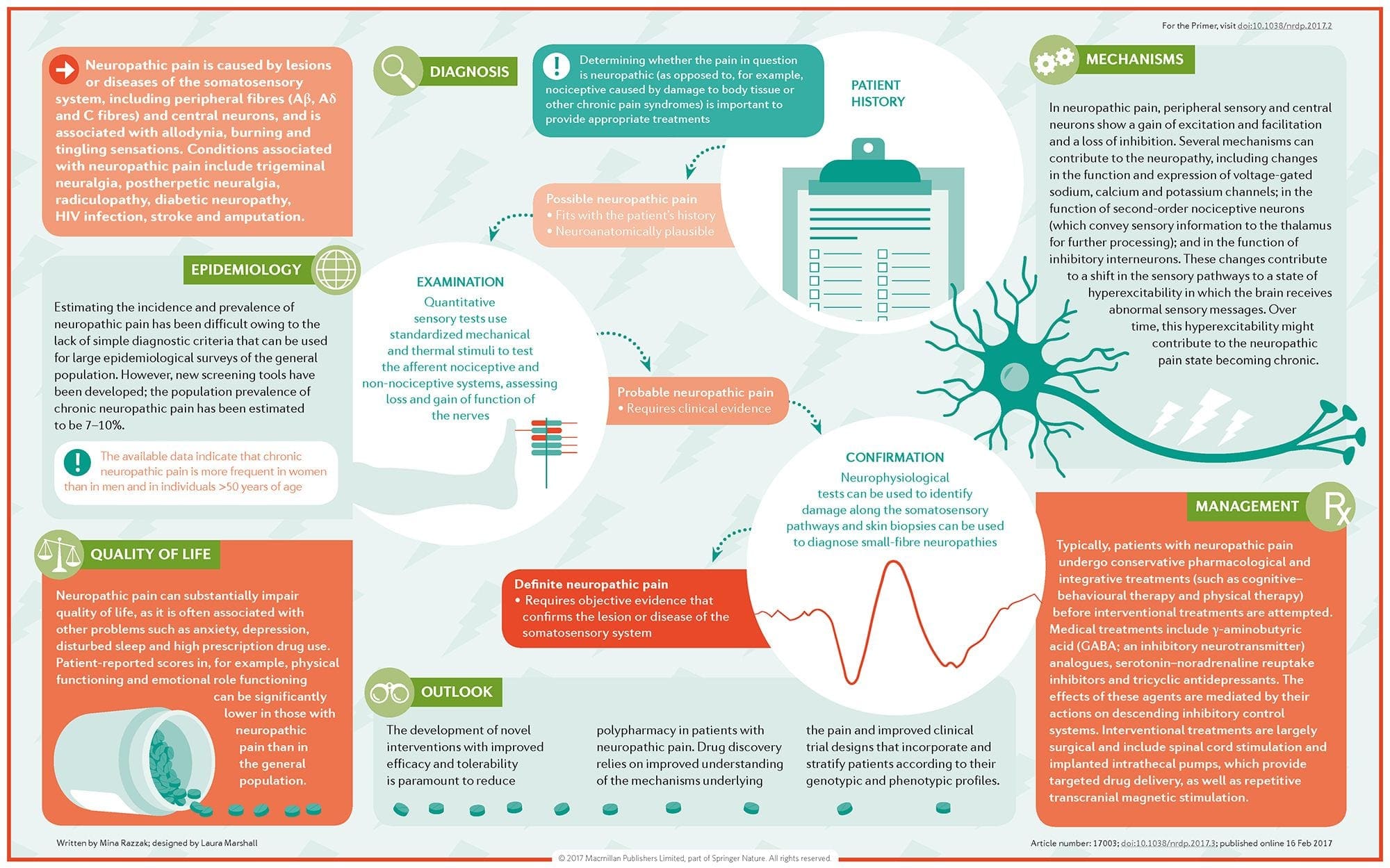




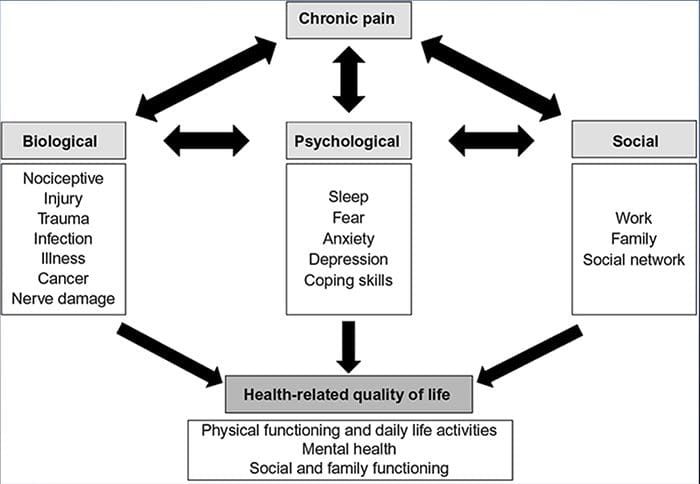
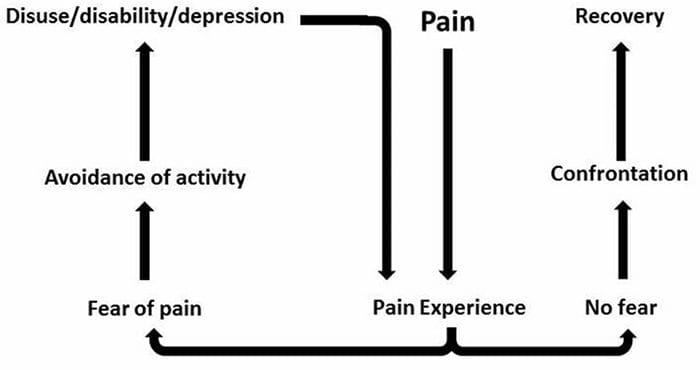
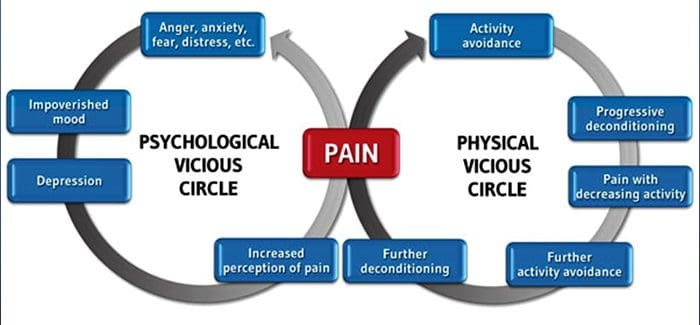
















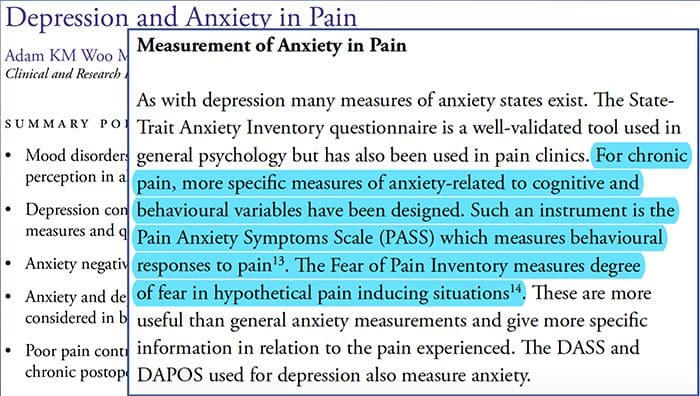


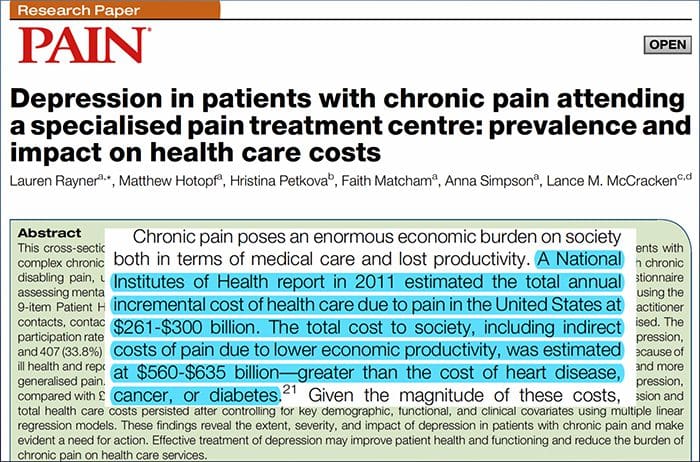



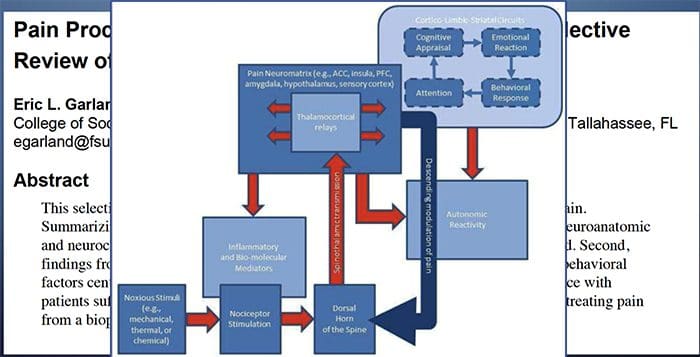






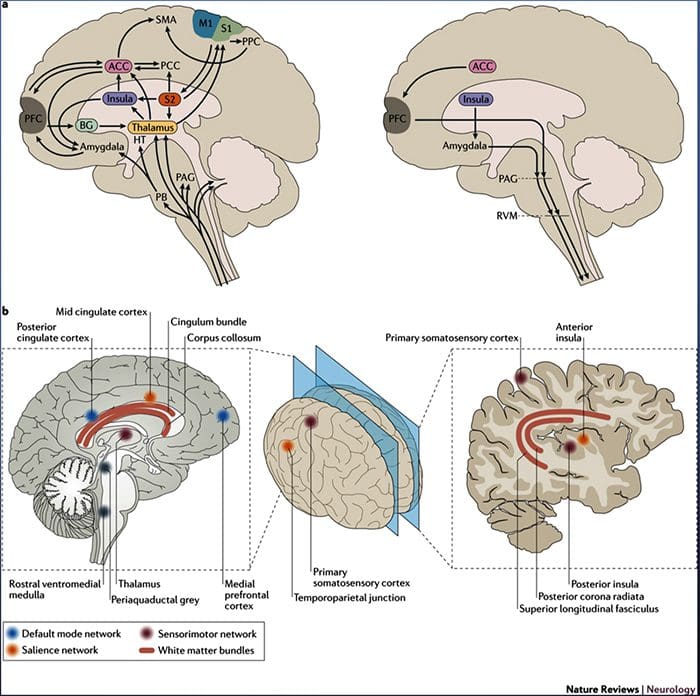

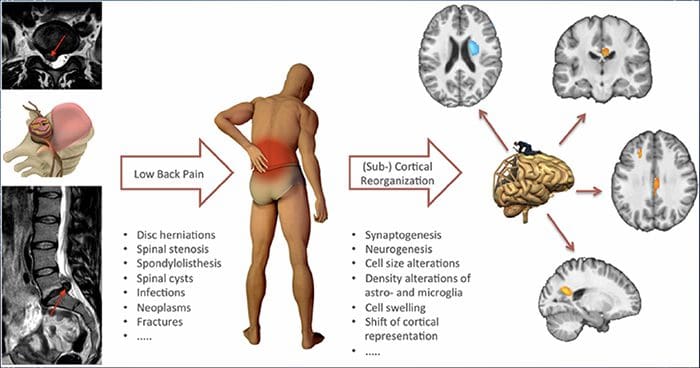

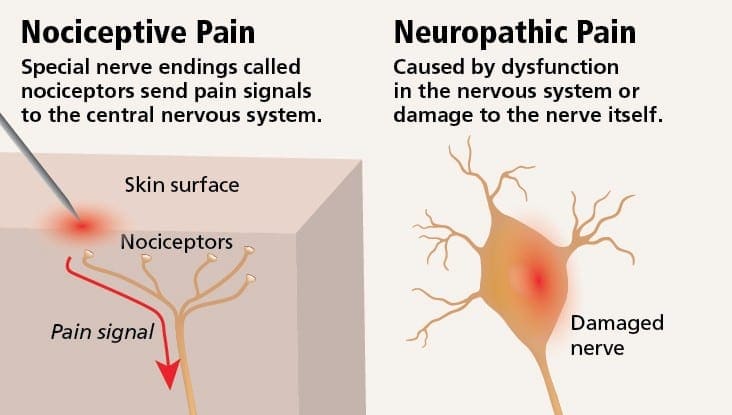
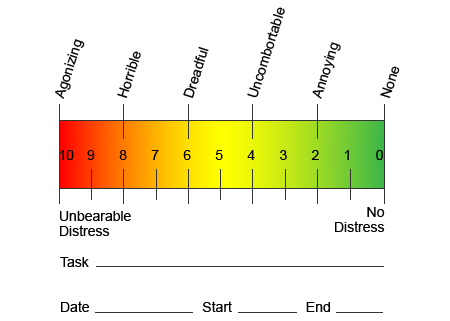


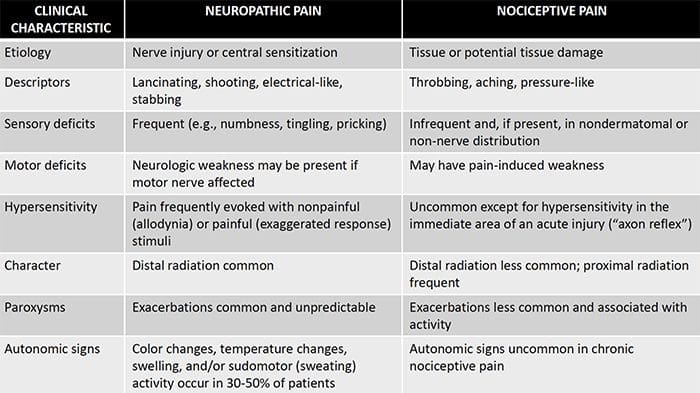
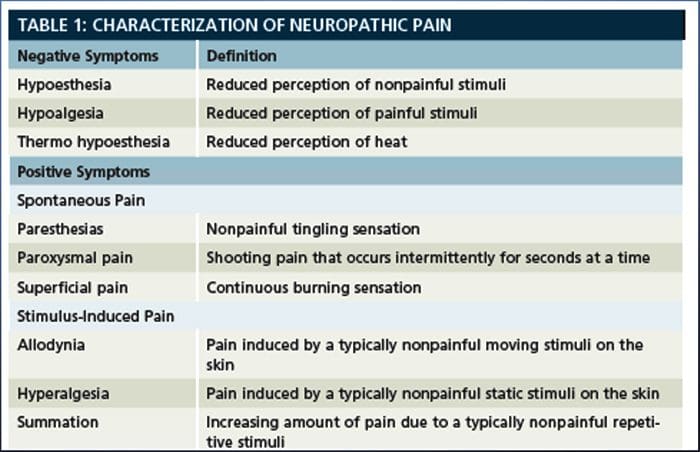
 پاتوژنز درد نروپاتیک
پاتوژنز درد نروپاتیک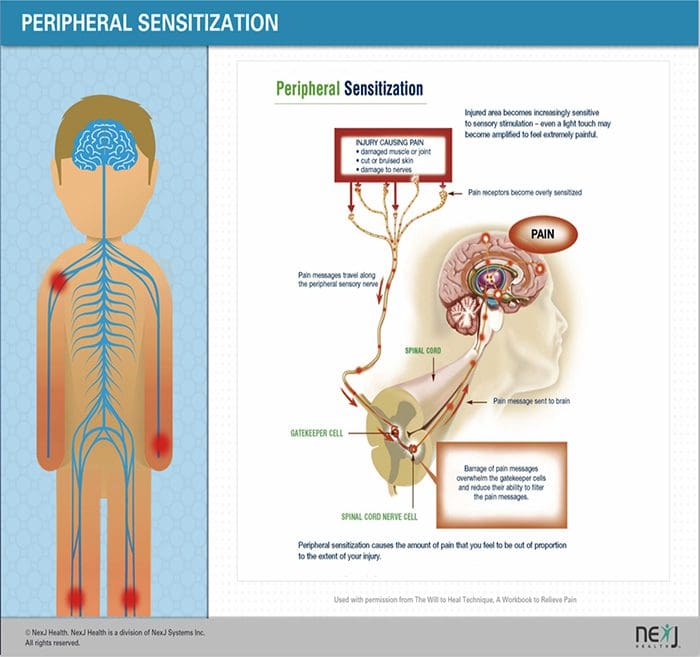
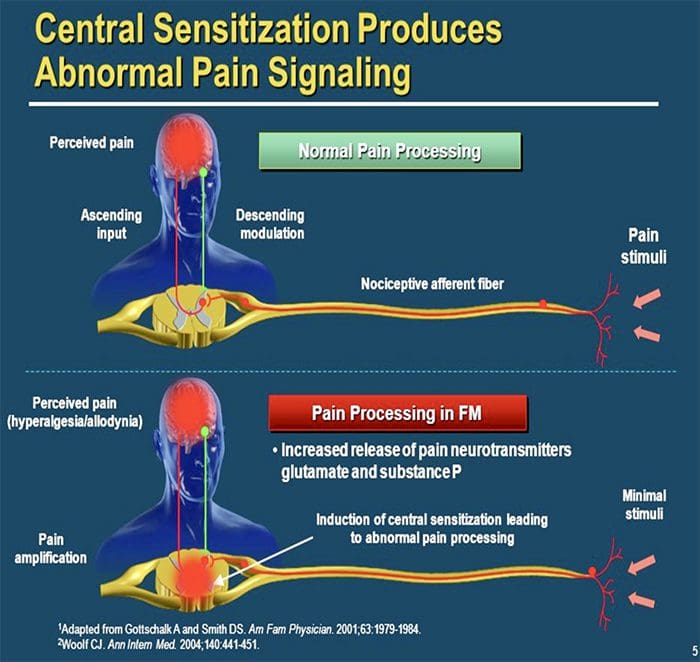
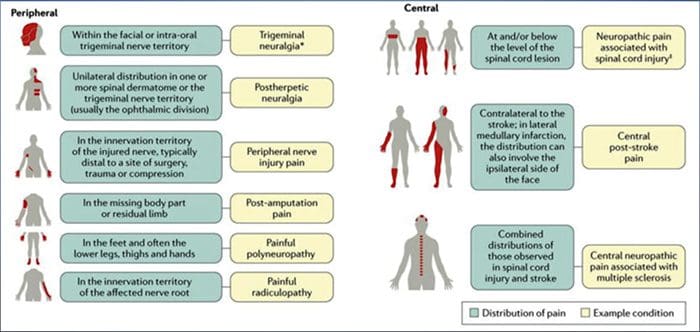 علل شایع
علل شایع
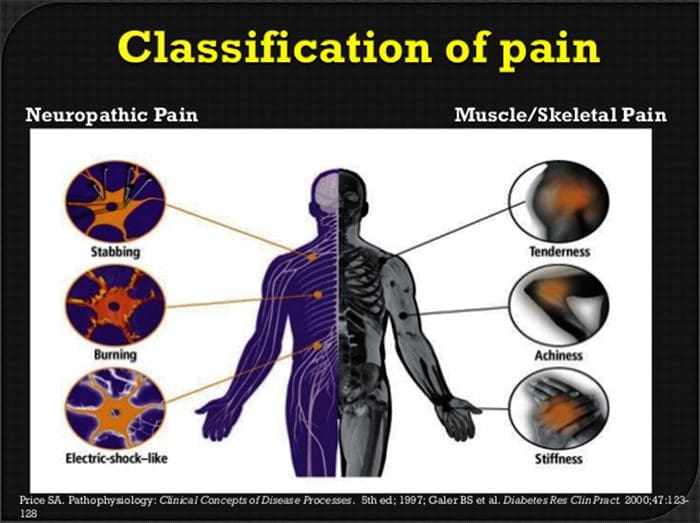
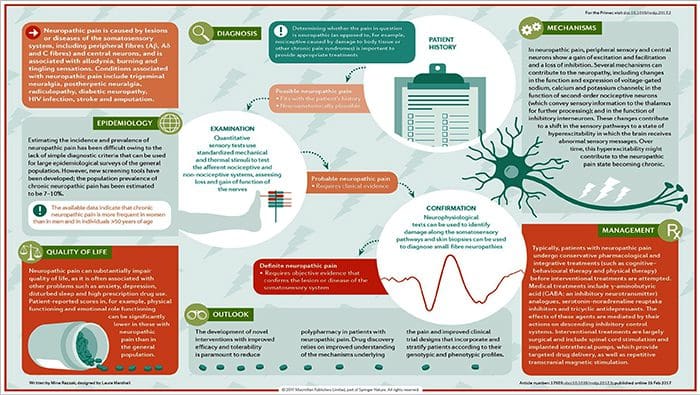
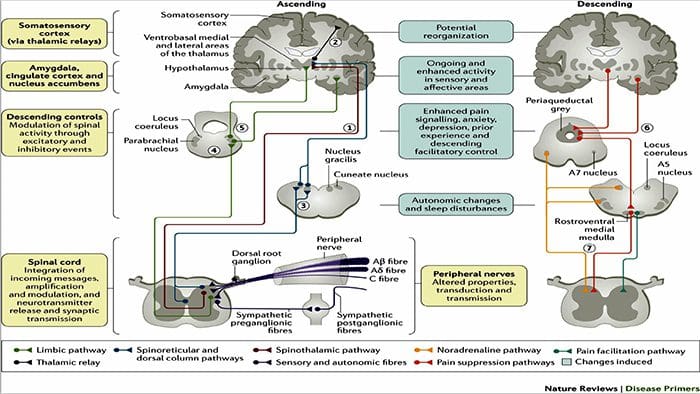
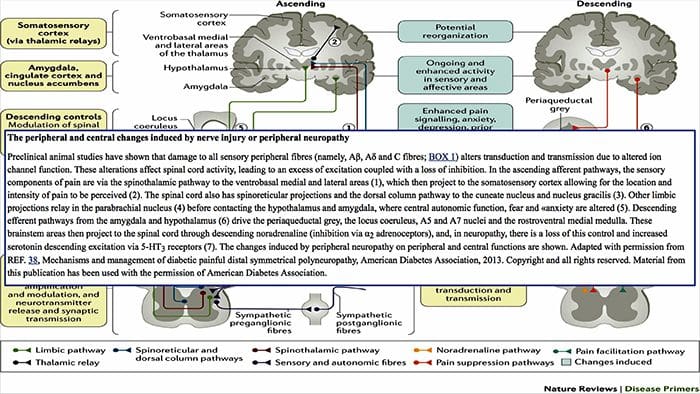

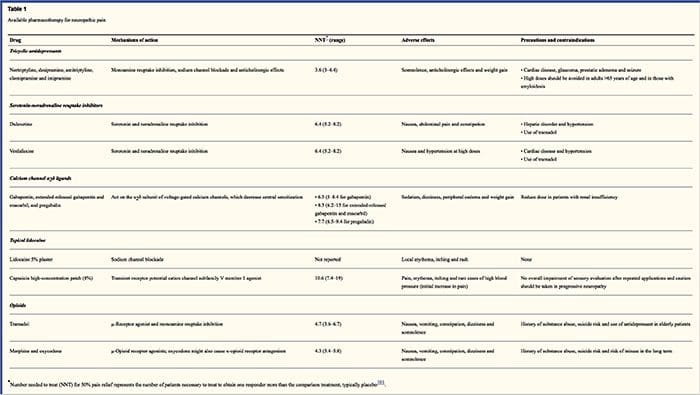




 درد اندام فانتوم و واقعیت افزوده
درد اندام فانتوم و واقعیت افزوده
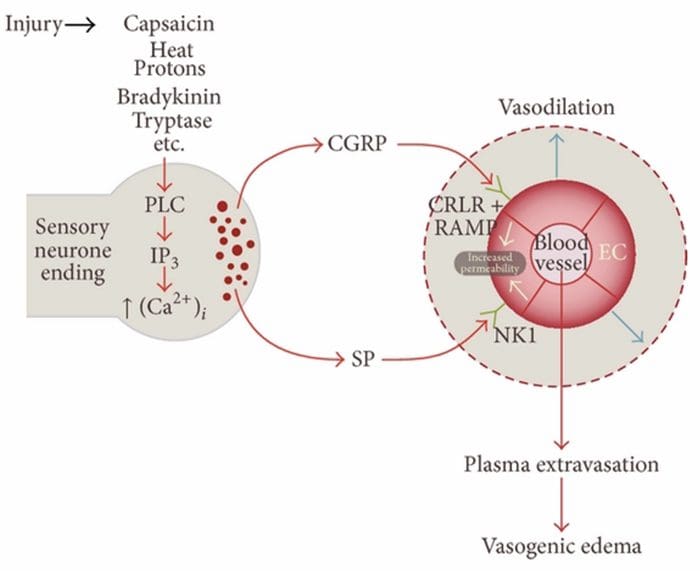 علل شایع
علل شایع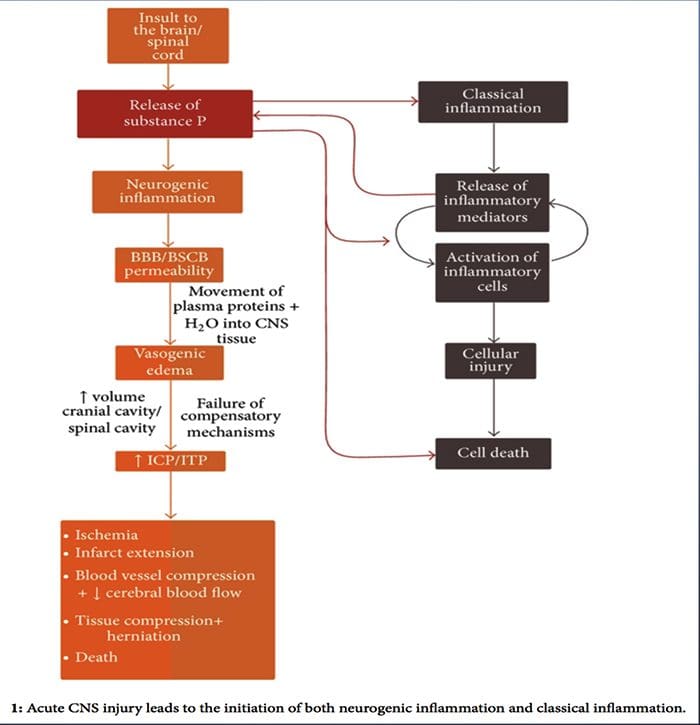


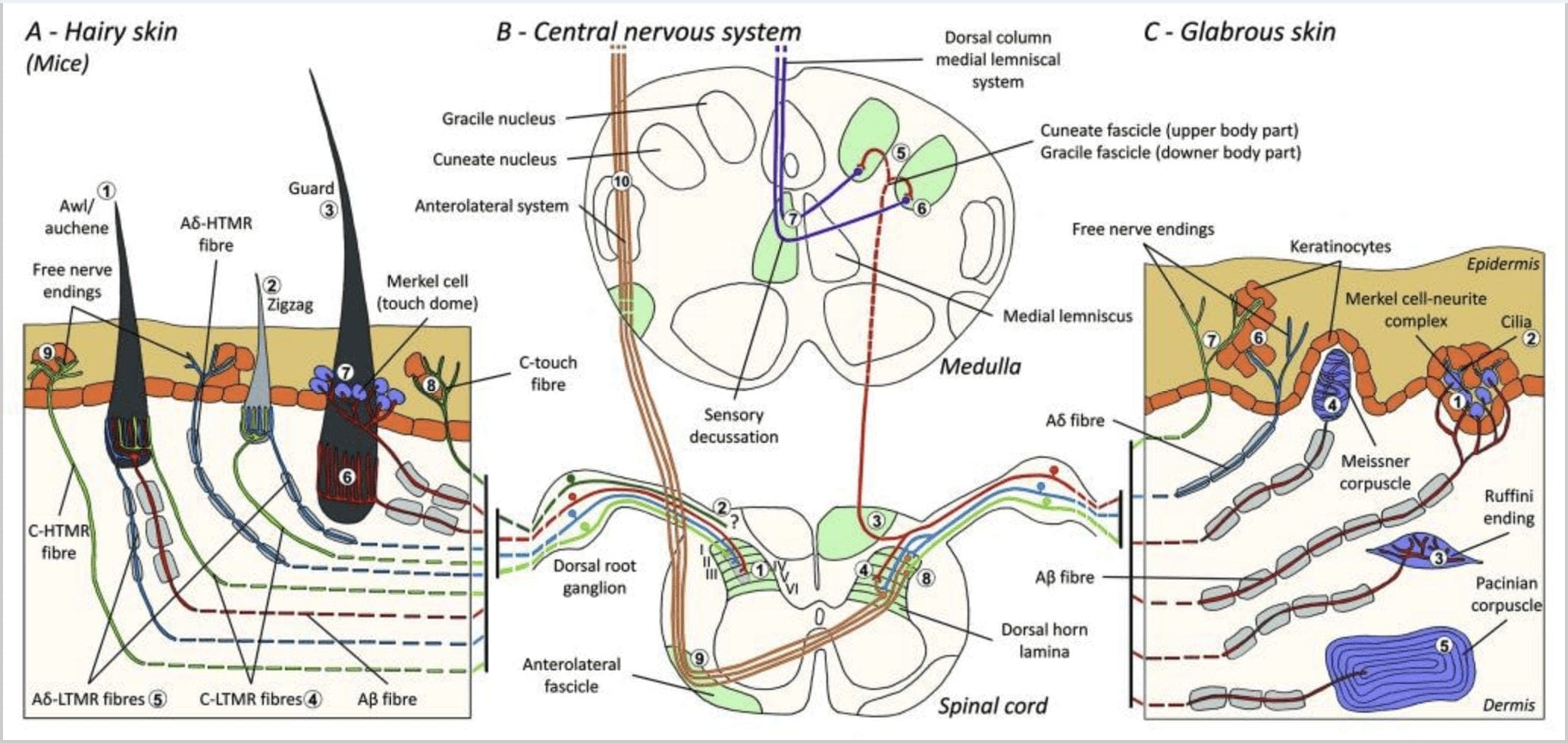
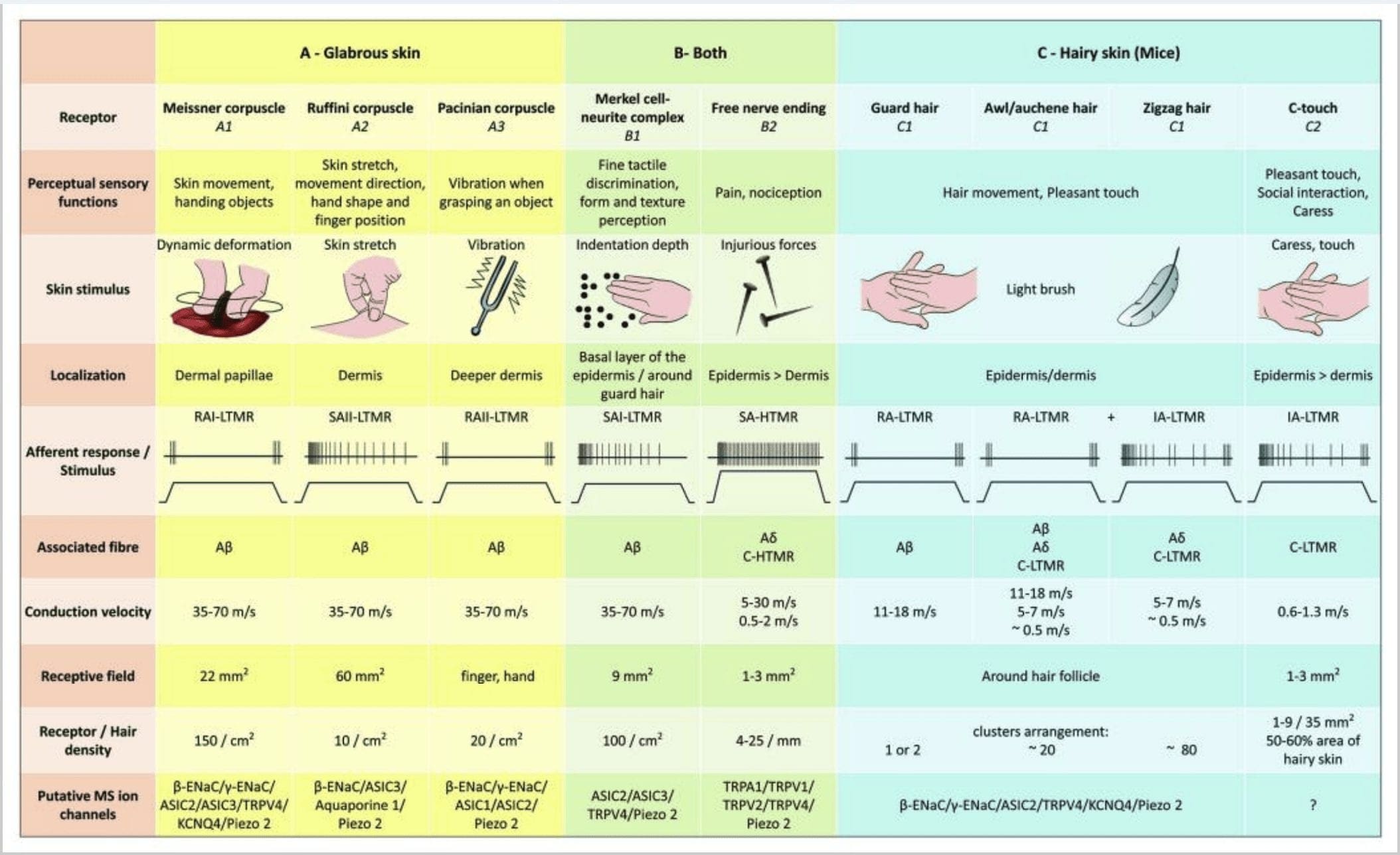

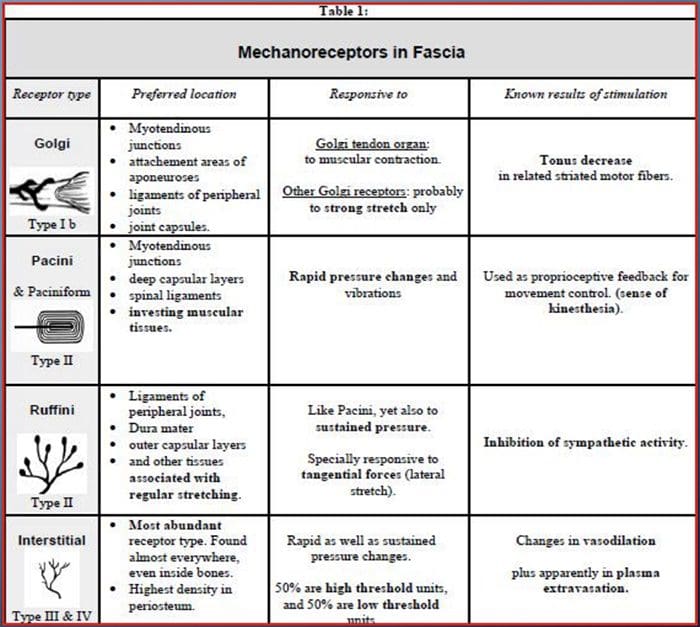 تئوری درد کنترل دروازه
تئوری درد کنترل دروازه
 مسیر لمنیسکال میانی ستون پشتی
مسیر لمنیسکال میانی ستون پشتی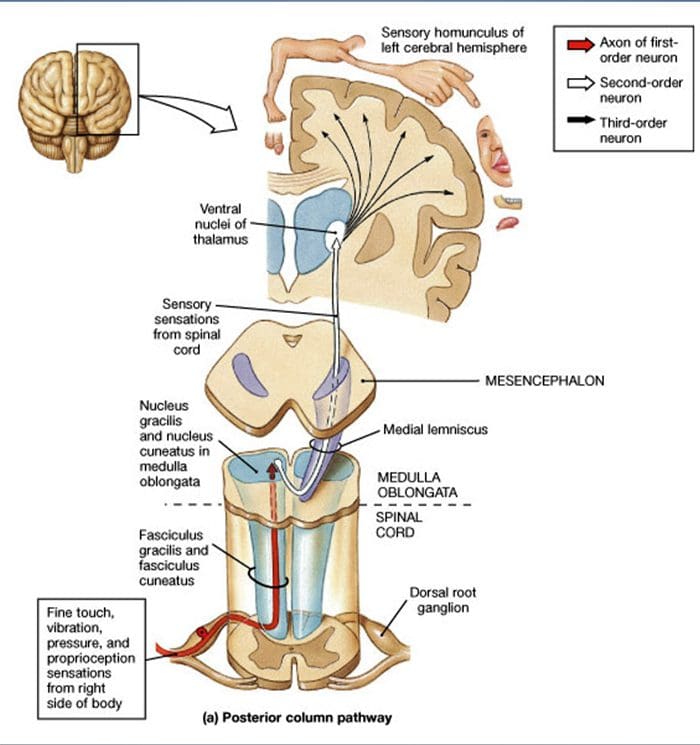 استفاده از گیرنده های مکانیکی محیطی برای تغییر ادراک درد
استفاده از گیرنده های مکانیکی محیطی برای تغییر ادراک درد
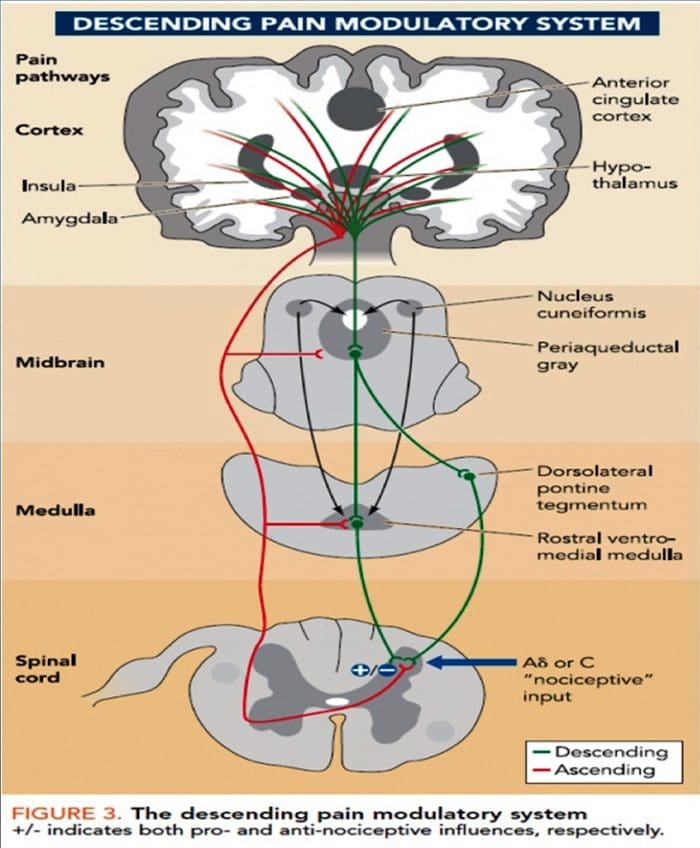
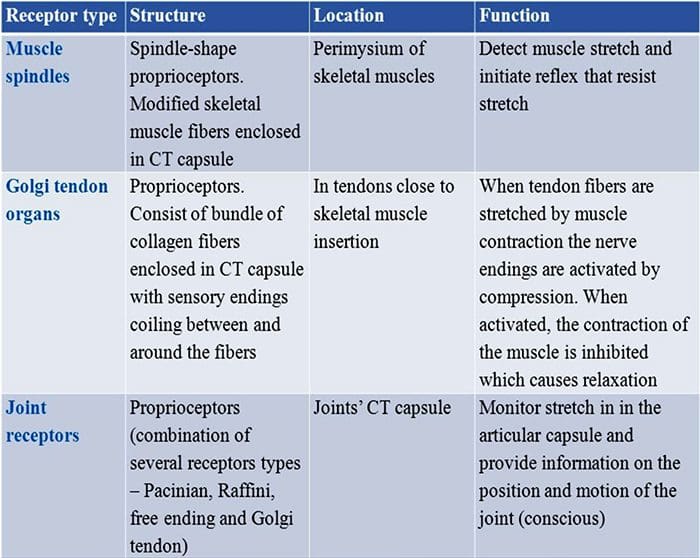
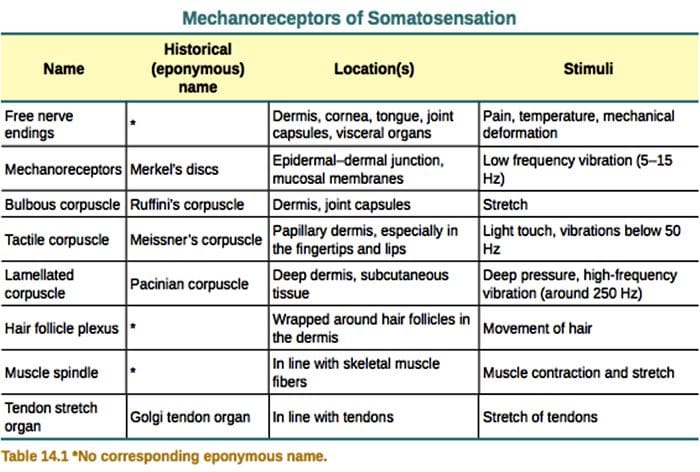 ارتعاش
ارتعاش
 لمس نور
لمس نور



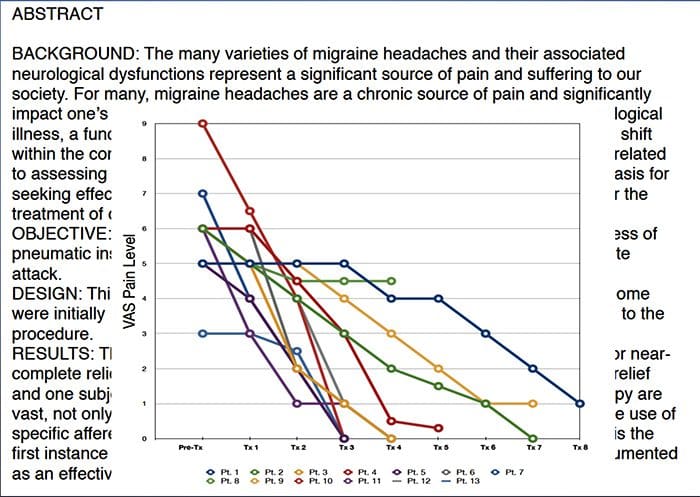

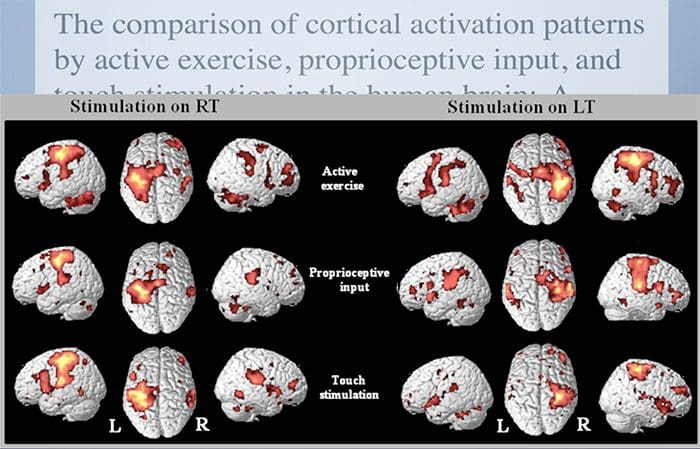
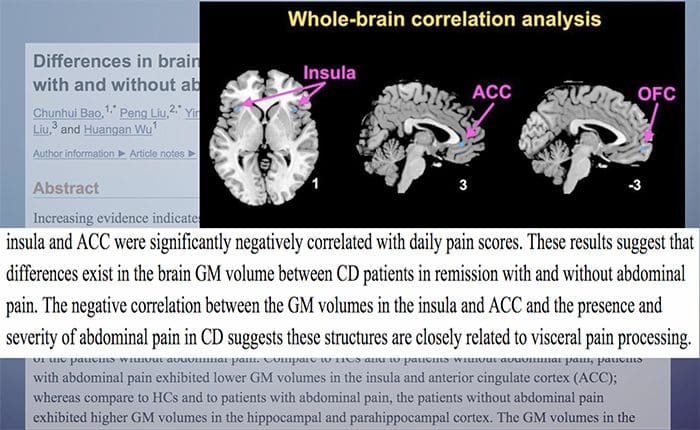

 مطالعه موردی
مطالعه موردی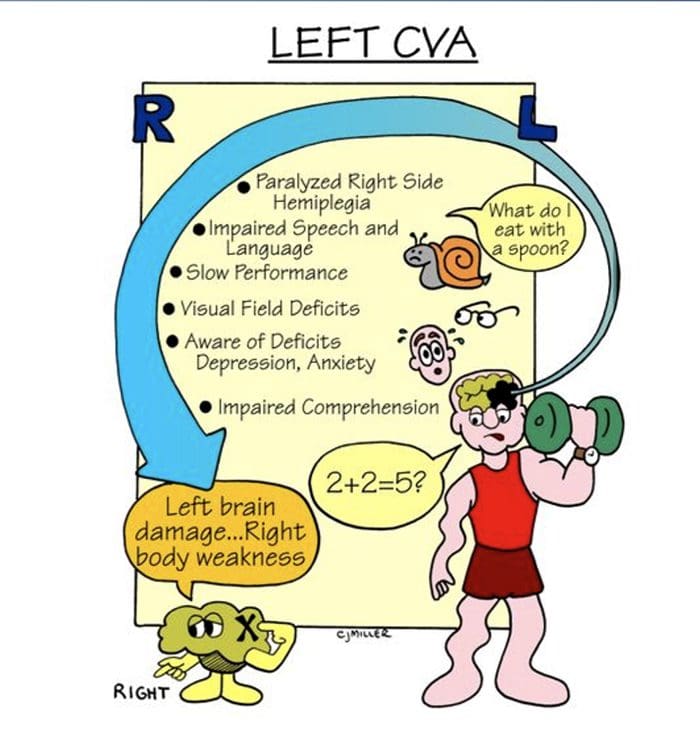 نکات برجسته معاینه فیزیکی
نکات برجسته معاینه فیزیکی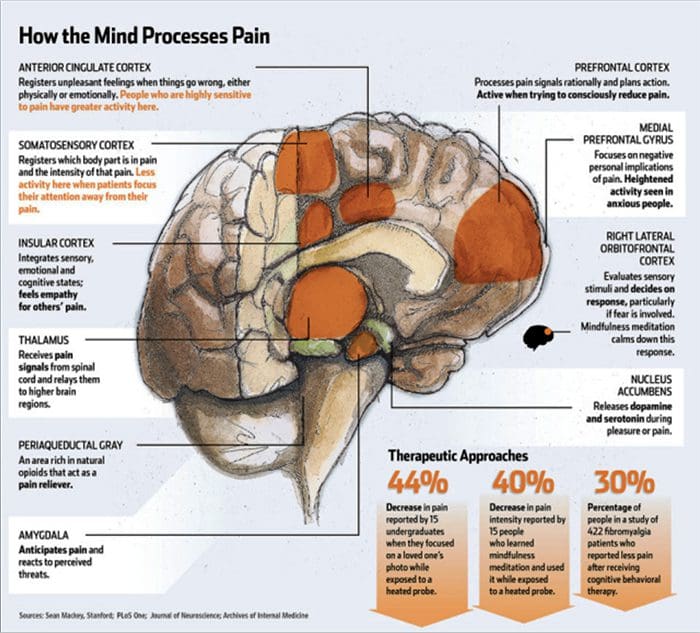 نکات برجسته معاینه فیزیکی
نکات برجسته معاینه فیزیکی
 بعد از دو روز
بعد از دو روز